Fruit morphology of the genus Pimpinella (Apiaceae) in Turkey
Yeter Yeşil 1,*, Emine Akalın 2, Aşkın Akpulat 3 & Cem Vural 4
1,2 Istanbul University, Faculty of Pharmacy, Deparment of Pharmaceutical Botany, 34116 Istanbul, Turkey.
3 Cumhuriyet University, Faculty of Education, Department of Biology Education, 58140 Sivas, Turkey.
4 Erciyes University, Faculty of Arts and Sciences, Department of Biology, 38039 Kayseri, Turkey.
* Author for correspondence: yeteryesil@yahoo.com, https://orcid.org/0000-0002-4458-7881
2 akaline@istanbul.edu.tr, https://orcid.org/0000-0002-0307-2128
3 aakpulat99@yahoo.com, https://orcid.org/0000-0001-8394-2746
4 vuralc@erciyes.edu.tr, https://orcid.org/0000-0001-9929-9935
| |
Abstract
Yeşil Y., Akalın E., Akpulat A. & Vural C. 2018. Fruit morphology of the genus Pimpinella (Apiaceae) in Turkey. Anales del Jardín Botánico de Madrid 75 (2): e072. https://doi.org/10.3989/ajbm.2509
To explore if fruit morphology could aid in taxonomy of the genus Pimpinella L., we have undertaken a study of fruits from 26 Turkish taxa of Pimpinella using light and scanning electron microscopy —SEM—. A great deal of inter and intraspecific variation for both fruit shape and surface was observed. Fruit shapes of Turkish taxa of Pimpinella range from oblong-cylindrical to subglobose and indumentum when present can be strigose, hispid and may include hamate trichomes. Variation in fruit surface is also considerable and allows recognizing nine different ornamentation patterns. However, variation in shape, surface ornamentation and indumentum is not tightly associated since species with similar fruit shapes do not necessarily have similar surface ornamentation. To jointly analyse fruit morphology together with the most commonly used morphological characters of the whole plant and to compare morphological evidence with available phylogenetic hypotheses, a cluster analysis was also performed: the Turkish species of Pimpinella were clustered into two distinct groups, the second one subdivided in another two subgroups.
Keywords. Apiaceae, morphology, Pimpinella, taxonomy, Turkey.
|
| |
Resumen
Title in Spanish: Morfología del fruto del género Pimpinella (Apiaceae) en Turquía.
Para comprobar el valor diagnóstico de la morfología del fruto en la taxonomía del género Pimpinella L., hemos estudiado los frutos de 26 táxones mediante microscopía óptica y electrónica de barrido —SEM—. Se ha observado una gran variabilidad inter e intraespecífica en la forma y la superficie del fruto. Las formas del fruto de los táxones turcos de Pimpinella varían de oblongo-cilíndricas a subglobosas, así como el fruto puede ser de estrigoso a híspido y tener a veces tricomas hamosos. La variabilidad de la superficie del fruto también es considerable y permite reconocer nueve patrones de ornamentación diferentes. Sin embargo, las variabilidades de la forma, la ornamentación de la superficie y el indumento no están estrechamente asociadas, ya que las especies con frutos de forma similar no necesariamente tienen una ornamentación similar. Para analizar conjuntamente la morfología del fruto y los caracteres morfológicos más comúnmente utilizados y para comparar la morfológica con las hipótesis filogenéticas disponibles, también se ha realizado un análisis de grupos: las especies turcas de Pimpinella formaron dos grupos y el segundo se subdividió en otros dos.
Palabras clave. Apiaceae, morfolog�a, Pimpinella, taxonom�a, Turqu�a.
|
INTRODUCTIONTOP
The c. 150 species constituting the genus Pimpinella L.―Apiaceae Lindl.―are distributed in temperate and subtropical regions of Eurasia and Africa including Madagascar. Turkey is one of the main centers of diversity for this genus along with Africa and Madagascar (Aksenov 1972; Abebe 1992). In his classical monograph, Wolff (1927) subdivided Pimpinella into three sections―P. sect. Reutera Boiss., P. sect. Tragium (Spreng.) DC. and P. sect. Tragoselinum (Mill.) DC.―based on petal color, fruit and petal indumentum, fruit ornamentation, and life form. Pimpinella sect. Reutera included species with yellow flowers and glabrous or hairy fruits; P. sect. Tragium included species mostly with white flowers and bristly or hairy, granular or tuberculate fruits, sometimes nearly glabrous or almost completely smooth; and P. sect. Tragoselinum included also white-flowered species with glabrous fruits. This classification has been widely adopted with some added characters. For instance, Pu & Watson (2005) in the Flora of China added calyx features. They recognized the first two of these sections: P. sect. Tragium including species with hairy or distinctly roughened fruits and obsolete calyx teeth, and P. sect. Tragoselinum, including species with glabrous fruits and obsolete or conspicuous calyx teeth. However, taxonomy of this genus, one of the most complex in the family, is relatively unsettled and phylogenetic studies have partly challenged Wolff’s sections (Magee & al. 2010). To achieve a stable taxonomy for this genus, it is not only necessary to refine the description of morphological characters as well as their patterns of variation and distribution across species but also to test those characters against solid molecular phylogenetic analyses.
For Turkey specifically, Matthews (1972) recognized 23 species with no infrageneric subdivison. Subsequent modifications to such treatment have involved transfers to, or from, other genera as well as new species. Pimpinella cruciata Bornm. & H.Wolff, which was identified as one of two varieties of P. anthriscoides Boiss. by Matthews in the Flora of Turkey, has been recently transferred to Tamamschjanella Pimenov & Kljuykov (Zakharova & al. 2012). Conversely, two names, treated under the genus Scaligeria DC. in the Flora of Turkey (Stevens 1972), have been confirmed to be part of Pimpinella, namely P. tripartita Kalen. and P. lazica (Boiss.) M.Hiroe (Hand 2011). In the same work P. affinis Ledeb. and P. squamosa Karjagin have been considered to be synonyms of P. peregrina L. and P. nudicaulis Trautv., respectively (Hand 2011). Finally, two new species have been recently described, P. ibradiensis Çingilbel & al. (Çingilbel & al. 2015) and P. enguezekensis Yıldırım & al. (Yeşil & al. 2016), so that currently Pimpinella includes 25 species―30 taxa, 8 of them endemic―in Turkey (Matthews 1972; Ertekin & Kaya 2005; Göktürk 2008; Menemen 2012; Çinbilgel & al. 2015; Yeşil & al. 2016).
Fruit characters are considered crucial in taxonomy throughout the whole Apiaceae as can be seen in any identification key (Engler 1927). The possibility of better characterizing fruits by using both anatomical characters and micromorphological features using SEM has stimulated numerous studies across the family in genera such as Bupleurum L. (Özcan 2004), Ferulago Koch (Akalın & Kızılarslan 2013), Ekimia H.Duman & M.F.Watson (Lyskov & al. 2015), Grammasciadium DC. (Bani & al. 2016a, 2016b) and Heracleum L. (Liu & Downie 2017).
In Pimpinella there have been several anatomical studies confined to important regions such as Iran (Khajepiri & al. 2010), Russia (Aksenov & Tikhomirov 1972), Africa and Madagascar (Magee & al. 2010). In a previous work, we conducted an anatomical study on the Turkish species (Akalın & al. 2016) that led to the recognition of four groups defined on the basis of fruit anatomical structure. Specifically, those four groups differed on the number and size of vallecular vitae, fruit shape, and trichomes and were partly compatible with the sections of Wollf (1927).
The taxonomic uncertainties together with the interest of this genus both at the taxonomic and phytochemical levels have prompted several molecular phylogenetic studies assessing relationships within Pimpinella. Tabanca & al. (2005) sampled 26 Turkish species of this genus focusing on distribution patterns of essential oils. Magee & al. (2010) attempted to elucidate the phylogenetic position of the African and Malagasy species but included 26 species from Eurasia in their analyses. Focusing on the genus circumscription, Fereidounfar & al. (2016) analyzed 52 Southwest Asian species of Pimpinella within a considerable sample of species from the family and concluded that P. sect. Reutera as well as Opsicarpium Mozaff. fall within Pimpinella and should be included in this genus. All the three studies were based on nuclear ribosomal ITS sequences and the first and third one also used plastid DNA sequences. Even though the focuses are different and sampling are not comprehensive, the phylogenetic positions of the species of Pimpinella included in two or more of these studies are to a large part consistent and thus there is some basis for phylogenetic relationships, which can be considered when taxonomic uncertainties are addressed. However, more research is needed on several fronts to clarify the taxonomy of this complex genus at a fine level.
The main purpose of this carpological study is to provide a detailed description of fruit morphology of 26 Turkish Pimpinella taxa―c. 87% of the Turkish taxa―including both micromorphological characters assessed using SEM, to contribute to species delimitation and infrageneric classification and to explore concordance with existing phylogenetic studies. We aim to aid in taxonomic classification by examining the fit of fruit characters with existing phylogenetic studies and by analyzing fruit variation together with the morphology of other organs.
MATERIAL AND METHODSTOP
Ripe fruits from Pimpinella corresponding to 26 taxa, 8 of them endemic, were obtained from specimens collected in different areas of Turkey (Table 1). Voucher specimens were deposited in ISTE―Herbarium of the Faculty of Pharmacy, Istanbul―. For the SEM micromorphological study, fruits were mounted on stubs using double adhesive tape and coated with gold-paladium. Specimens were examined under a JEOL Neoscope 5000 electron microscope at 10.00 kV. Macromorphological observations were made, and photograps were taken, with a LEICA DFC 295 stereo microscope with a digital camera. Measurements of mericarps, using LEICA software, were performed on at least five mature fruits from each of the 26 studied taxa. The main morphological features recorded are summarized in Table 2. For descriptions and terminology of our micromorphological observations, we follow Özcan (2004), Bani & al. (2016a, 2016b) and Liu & Downie (2017). Overall shapes of mericarps were classified according to Botanical Latin (Stearn 2005) and Aksenov & al. (1972). In addition, to explore phenetic similarity among the Pimpinella taxa, we performed a cluster analyses. Specifically a hierarchical agglomerative clustering analysis―method:ward.D―using the hclust function in R package v3.3.1. (R Development Core Team 2018) was run to construct a dendrogram. For this, the overlapping characteristics were previously eliminated (Wolf 1927; Abebe 1992) and catergorical variables were trasnformed into binary. Twenty-eight binary characters―presence/absence―from the fruits and from other plant organs were included in the analysis. Fruit characters are size, shape, indumentum (Table 2) and the micromorphological ones described below under results. Morphological characters from other plant organs are flower color―white, yellow, pink, red―, fruit indumentum―hairy or glabrous―, leaf shape―simple or pinnate―, and bracts and bracteoles―presence/absence.
Table 1. The list and collection numbers of studied Turkish taxa of Pimpinella L.
| Taxon |
Grid City |
Location |
Altitude m a.s.l. |
Collection Number |
| P. affinis Ledeb. |
B7 Erzincan |
Kemaliye, Sançiçek Plateau, 13–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
1790 |
ISTE 96851 |
| P. anisetum Boiss. & Balansa |
B7 Erzincan |
Spikor mountain, Çayırlı road, 10 km after Erzincan, 14–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
2293 |
ISTE 95807 |
| P. anisum L. |
A9 Ardahan |
Kutul, Yalnızçam Forest, 4–IX–2010, A. Akpulat s.n. leg. |
800 |
ISTE 96842 |
| P. aromatica M.Bieb. |
B7 Erzincan |
Spikor mountain, Kolgeçmez pass, 14–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
2360 |
ISTE 94693 |
| P. aurea DC. |
C10 Hakkâri |
6 km after Hakkâri-Yüksekova turnout, 27–VII–2012, E. Akalın and U. Uruşak s.n. leg. |
2185 |
ISTE 98881 |
| P. cappadocica Boiss. & Balansa var. cappadocica |
B7 Sivas |
Mut-Kırobası, 7 km from Mut, 30–VI–2012, A. Akpulat 4810 leg. |
514 |
ISTE 10117 |
| P. corymbosa Boiss. |
B7 Erzincan |
Spikor Mountain, Çayırlı road, 24 km from Erzincan, 14–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
1318 |
ISTE 95805 |
| P. cretica Poir. var. cretica |
C1 Aydın |
Priene ancient city, left side of entrance, 4–VI–2012, E. Akalın and U. Uruşak s.n. leg. |
26 |
ISTE 98669 |
| P. eriocarpa Banks & Sol. |
B7 şanlıurfa |
Northwest of Korukezen village, 6–XI–2012, E. Akalın and U. Uruşak s.n. leg. |
840 |
ISTE 98778 |
| P. enguezekensis Yıldırım & al. |
B6 Malatya |
Darende District, Ergü road, Kilise location, 22–VII–2015, H.Yıldırım HY3492 leg. |
1420 |
ISTE 107588 |
| P. flabellifolia (Boiss.) Benth. & Hook. ex Drude |
B6 Sivas |
Divriği, Arguvan-Divriği road, between Beldibi-Yeşilyol villages, 21–VII–2015, H. Yıldırım HY3472 leg. |
1451 |
ISTE 107580 |
| P. ibradiensis Çinbilgel & al. |
C3 Antalya |
İbradı, Toka Yayla, 2–VII–2011, Çinbilgel 7975 and Eren leg. |
1527 |
ISTE 115057 |
| P. isaurica V.A.Matthews subsp. isaurica |
C4 Konya |
Ermenek, around Keben fountain, 28–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
1293 |
ISTE 95813 |
| P. kotschyana Boiss. |
B1 Manisa |
Spil Mountain, Spil roadside, 5–VII–2011, E. Akalın and U. Uruşak s.n. leg. |
306 |
ISTE 95735 |
| P. lazica (Boiss.) M.Hiroe |
A8 Rize |
Çamlıhemşin, Boğaziçi village, Tunuslu town, 6–IX–2010, A. Akpulat and M. Tekin 16 leg. |
600 |
ISTE 96846 |
| P. nephrophylla Rech.f. & Riedl |
B8 Diyarbakır |
Eğil, Eğil castle, 13–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
900 |
ISTE 95784 |
| P. nudicaulis Trautv. |
B7 Erzincan |
Tercan, Gahmut Plateau, 10–VIII–2009, E. Akalın and U. Uruşak s.n. leg. |
1910 |
ISTE 101345 |
| P. oliverioides Boiss. & Hausskn. |
B9 Van |
Van-Hoşap, Güzeldere pass, 18–VIII–1993, Y. Altan 5552 leg. |
2800 |
GAZI |
| P. paucidentata V.A.Matthews |
B6 Malatya |
Darende, Ağılbaşı town, Ergü road, Kilise location, 10–VIII–2017, Y. Yeşil s.n. leg. |
1420 |
ISTE 115020 |
| P. peregrina L. |
B1 Manisa |
Hatipler-şatırlar, Hatipler village, 6–VII–2011, E. Akalın and U. Uruşak s.n. leg. |
288 |
ISTE 95775 |
| P. peucedanifolia Fisch. |
B7 Erzincan |
Spikor mountain, Mecidiye location, 13–VIII–2010, E. Akalın and U. Uruşak s.n. leg. |
2310 |
ISTE 94695 |
| P. puberula (DC.) Boiss. |
C9 Hakkâri |
Hakkâri-Van, 12 km after Hakkâri, 27–VII–2012, E. Akalın and U. Uruşak s.n. leg. |
1446 |
ISTE 98878 |
| P. rhodantha Boiss. |
A9 Ardahan |
Çataldere Plateau, 27–VII–2011, B. Gürdal and S. Esen s.n. leg. |
1548 |
ISTE 97267 |
| P. saxifraga L. |
A6 Ordu |
Koyulhisar-Mesudiye, 11–VIII–2010, E. Akalın and U. Uruşak s.n. leg. |
1370 |
ISTE 94675 |
| P. sintenisii H.Wolff |
C8 Mardin |
Darulzaferan Monastery, 11–VI–2012, E. Akalın and U. Uruşak s.n. leg. |
1212 |
ISTE 98789 |
| P. tragium subsp. pseudotragium (DC.) V.A.Matthews |
B7 Erzincan |
Spikor mountain, Kolgeçmez pass, 14–VIII–2011, E. Akalın and U. Uruşak s.n. leg. |
2684 |
ISTE 95811 |
Table 2. Fruit measurements and features of the mericarps of Turkish taxa of Pimpinella L.
| Taxon |
Fruit length and width (mm) |
Length/width ratio |
Indumentum |
Shape of fruit |
| P. affinis Ledeb. |
1.65–1.75 × 0.77–0.8 |
2.06 |
pubescens |
elliptic |
| P. anisetum Boiss. & Balansa |
1.5–1.57 × 1–1.08 |
1.5 |
strigose |
ovoid |
| P. anisum L. |
3.9–4 × 1.5–1.53 |
2.6 |
strigose |
ovoid-subglobose |
| P. aromatica M.Bieb. |
1.85–1.9 × 1.21–1.25 |
1.52 |
strigose |
ovoid |
| P. aurea DC. |
2.23–2.25 × 1.58–1.6 |
1.40 |
pubescens |
subglobose |
| P. cappadocica Boiss. & Balansa var. cappadocica |
1.97–2 × 1.14–1.15 |
1.73 |
hispid |
ovoid |
| P. corymbosa Boiss. |
1.9–2 × 1.95–1 |
2 |
pubescens |
ovoid-subglobose |
| P. cretica Poir. var. cretica |
1.42–1.5 × 0.95–1 |
1.5 |
strigose |
ovoid-globose |
| P. enguezekensis Yıldırım & al. |
2.57–2.6 × 1.69–1.7 |
1.52 |
glabrous |
oblong-ovoid |
| P. eriocarpa Banks & Sol. |
1.57–1.6 × 0.78–0.8 |
2 |
hispid-subhamate |
elliptic |
| P. flabellifolia (Boiss.) Benth. & Hook. ex Drude |
3.9–4 × 2.25–2.3 |
1.73 |
rarely hispid |
oblong-ovoid |
| P. ibradiensis Çinbilgel & al. |
4–5.5 × 1–2 |
2.89 |
glabrous |
oblong-cylindrical |
| P. isaurica V.A.Matthews subsp. isaurica |
3.4–3.5 × 0.95–0.98 |
3.57 |
hirsute |
oblong-cylindrical |
| P. kotschyana Boiss. |
2.6–2.65 × 1.18–1.2 |
2.20 |
hispid |
ovoid-subglobose |
| P. lazica (Boiss.) M.Hiroe |
2.7–2.8 × 1.6–1.66 |
1.68 |
glabrous |
oblong-ovoid |
| P. nephrophylla Rech.f. & Riedl |
2.3–2.35 × 0.85–0.87 |
2.70 |
glabrous |
oblong |
| P. nudicaulis Trautv. |
3.6–3.65 ×1.3–1.35 |
2.70 |
glabrous |
oblong-cylindrical |
| P. oliverioides Boiss. & Hausskn. |
4.25–4.35 × 1.6–1.64 |
2.65 |
pubescens |
oblong |
| P. peregrina L. |
1.9–2 × 0.9–0.94 |
2.11 |
hispid |
eliptic |
| P. peucedanifolia Fisch. |
2.55–2.6 × 0.58–0,6 |
4.33 |
glabrous |
oblong-cylindrical |
| P. paucidentata V.A.Matthews |
2.15–2.2 × 0.7–0.71 |
3.07 |
glabrous |
oblong |
| P. puberula (DC.) Boiss. |
1.6–1.67 × 1.05–1.10 |
1.52 |
hamate |
ovoid-globose |
| P. rhodantha Boiss. |
2.9–3.1 × 1.8–1.92 |
1.61 |
glabrous |
oblong-ovoid |
| P. saxifraga L. |
2.1–2.2 × 1.7–1.78 |
1.23 |
glabrous |
oblong-ovoid |
| P. sintenisii |
1.8–1.95 × 0.65–0.7 |
2.76 |
glabrous |
oblong |
| P. tragium subsp. pseudotragium |
2.3–2.42 × 1.4–1.47 |
1.64 |
hamate |
oblong-ovoid |
RESULTSTOP
Macromorphology of fruitsTOP
Fruit shape of Turkish taxa of Pimpinella can be referred to the following categories: oblong-cylindrical, oblong, elliptic, ovoid-subglobose, oblong-ovoid, ovoid, ovoid-globose, and subglobose (Fig. 1). The ratio of fruit length to width varies between 4.33 and 1.5. The largest fruits―3.4–5.5 mm long―are found in P. ibradiensis―light microscopy photo not shown―, P. oliveroides Boiss. & Hausskn., P. nudicaulis, P. anisum L., P. isaurica V.A.Matthews subsp. isaurica, and P. flabellifolia (Boiss.) Benth. & Hook. ex Drude, whereas the smallest―1.42–1.5 mm―is found in P. cretica Poir. var. cretica (Table 2). Fruit indumentum has been assigned to the following states: pubescens, strigose, rarely hispid, hispid, hamate, hamate or glabrous. Tichome surface is always verrucate.
|
 Fig. 1. Mericarps of the Turkish taxa of Pimpinella L.: a, P. nudicaulis Trautv.; b, P. isaurica V.A.Matthews subsp. isaurica; c, P. peucedanifolia Fisch.; d, P. oliverioides Boiss. & Hausskn.; e, P. nephrophylla Rech.f. & Riedl; f, P. sintenisii H.Wolff; g, P. paucidentata V.A.Matthews; h, P. eriocarpa Banks & Sol.; i, P. peregrina L.; j, P. affinis Ledeb.; k, P. anisum L.; l, P. kotschyana Boiss.; m, P. corymbosa Boiss.; n, P. flabellifolia (Boiss.) Benth. & Hook. ex Drude; o, P. rhodantha Boiss.; p, P. enguezekensis Yıldırım & al.; q, P. lazica (Boiss.) M.Hiroe; r, P. tragium subsp. pseudotragium (DC.) V.A.Matthews; s, P. saxifraga L.; t, P. cappadocica Boiss. & Balansa var. cappadocica; u, P. aromatica M.Bieb.; v, P. anisetum Boiss. & Balansa; w, P. puberula (DC.) Boiss.; x, P. cretica Poir. var. cretica; y, P. aurea DC. Scale bar: 1 mm. Fig. 1. Mericarps of the Turkish taxa of Pimpinella L.: a, P. nudicaulis Trautv.; b, P. isaurica V.A.Matthews subsp. isaurica; c, P. peucedanifolia Fisch.; d, P. oliverioides Boiss. & Hausskn.; e, P. nephrophylla Rech.f. & Riedl; f, P. sintenisii H.Wolff; g, P. paucidentata V.A.Matthews; h, P. eriocarpa Banks & Sol.; i, P. peregrina L.; j, P. affinis Ledeb.; k, P. anisum L.; l, P. kotschyana Boiss.; m, P. corymbosa Boiss.; n, P. flabellifolia (Boiss.) Benth. & Hook. ex Drude; o, P. rhodantha Boiss.; p, P. enguezekensis Yıldırım & al.; q, P. lazica (Boiss.) M.Hiroe; r, P. tragium subsp. pseudotragium (DC.) V.A.Matthews; s, P. saxifraga L.; t, P. cappadocica Boiss. & Balansa var. cappadocica; u, P. aromatica M.Bieb.; v, P. anisetum Boiss. & Balansa; w, P. puberula (DC.) Boiss.; x, P. cretica Poir. var. cretica; y, P. aurea DC. Scale bar: 1 mm.
|
|
Micromorphology of fruit surfaceTOP
The mericarp surface shows a variety of micromorphological patterns at the SEM (figs. 2–4). The following nine types of ornamentation were observed in this study:
|
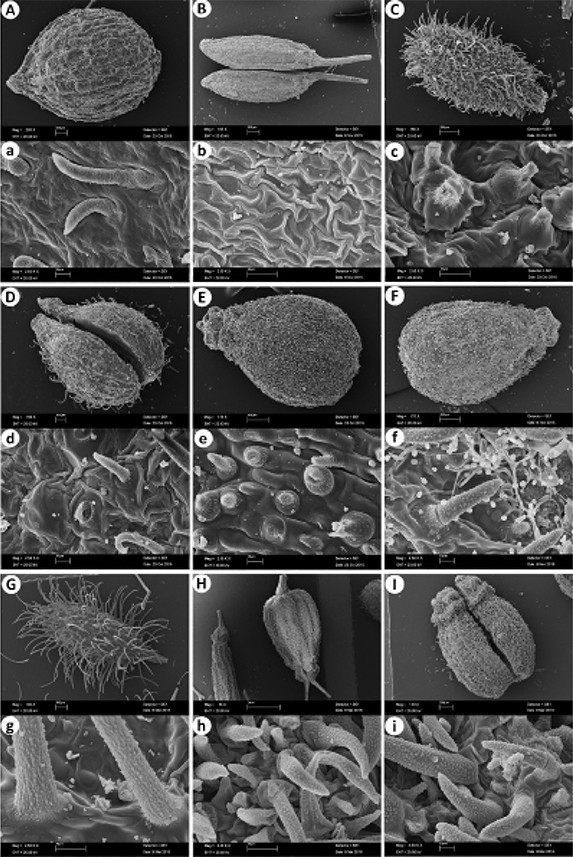 Fig. 2. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. cretica Poir.; B, b, P. nephrophylla Rech.f. & Riedl; C, c, P. peregrina L.; D, d, P. puberula (DC.) Boiss.; E, e, P. anisetum Boiss. & Balansa; F, f, P. aromatica M.Bieb.; G, g, P. eriocarpa Banks & Sol.; H, h, P. anisum L.; I, i, P. corymbosa Boiss. Fig. 2. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. cretica Poir.; B, b, P. nephrophylla Rech.f. & Riedl; C, c, P. peregrina L.; D, d, P. puberula (DC.) Boiss.; E, e, P. anisetum Boiss. & Balansa; F, f, P. aromatica M.Bieb.; G, g, P. eriocarpa Banks & Sol.; H, h, P. anisum L.; I, i, P. corymbosa Boiss.
|
|
|
 Fig. 3. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. nudicaulis Trautv.; B, b, P. peucedanifolia Fisch. ex Ledeb.; C, c, P. affinis Ledeb.; D, d, P. lazica (Boiss.) M.Hiroe; E, e, P. saxifraga L.; F, f, P. sintenisii H.Wolff; G, g, P. rhodantha Boiss.; H, h, P. enguezekensis Yıldırım & al.; I, i, P. isaurica V.A.Matthews subsp. isaurica. Fig. 3. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. nudicaulis Trautv.; B, b, P. peucedanifolia Fisch. ex Ledeb.; C, c, P. affinis Ledeb.; D, d, P. lazica (Boiss.) M.Hiroe; E, e, P. saxifraga L.; F, f, P. sintenisii H.Wolff; G, g, P. rhodantha Boiss.; H, h, P. enguezekensis Yıldırım & al.; I, i, P. isaurica V.A.Matthews subsp. isaurica.
|
|
|
 Fig. 4. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. aurea DC.; B, b, P. cappadocica Boiss. & Balansa var. cappadocica; C, c, P. flabellifolia (Boiss.) Benth. & Hook. ex Drude; D, d, P. kotschyana Boiss.; E, e, P oliverioides Boiss. & Hausskn. ex Boiss.; F, f, P. tragium subsp. pseudotragium (DC.) V.A.Matthews; G, g, P. ibradiensis Çingilbel & al. Fig. 4. SEM micrographs of mericarp coat surface in the genus Pimpinella L.: A, a, P. aurea DC.; B, b, P. cappadocica Boiss. & Balansa var. cappadocica; C, c, P. flabellifolia (Boiss.) Benth. & Hook. ex Drude; D, d, P. kotschyana Boiss.; E, e, P oliverioides Boiss. & Hausskn. ex Boiss.; F, f, P. tragium subsp. pseudotragium (DC.) V.A.Matthews; G, g, P. ibradiensis Çingilbel & al.
|
|
Type 1, smooth-rugose: among the Turkish species, this distinct surface ornamentation pattern is only found in P. cretica var. cretica. The mericarp surface is covered by strigose hairs (fig. 2A, a).
Type 2, rugose: irregularly colliculate with interconnected foldings. It occurs on P. nephrophylla Rech.f. & Riedl, P. peregrina, and P. puberula (DC.) Boiss. (fig. 2).
Type 3, rugulose: colliculate-tuberculate surface pattern, but with very few tubercules―P. anisetum Boiss. & Balansa, P. aromatica M.Bieb., and P. eriocarpa Banks & Sol.―(fig. 2).
Type 4, rugose-striate: with uneven, short and incomplete folds bearing secondary striate parallel furrows―P. anisum, P. corymbosa Boiss., P. nudicaulis, and P. peucedanifolia Fisch. ex Ledeb.―(figs. 2–3).
Type 5, rugose-reticulate: with nerve-like elevations that come from a reticular surface―P. affinis, P. lazica, P. saxifraga L., and P. sintenisii H.Wolff―(fig. 3).
Type 6, reticulate-striate: striate with longitudinal folds―P. rhodantha Boiss. and P. enguezekensis― (fig. 3).
Type 7, striate-ruminate: densely striate with irregular folds―P. isaurica subsp. isaurica― (fig. 3I, i).
Type 8, striate: irregularly colliculate and with folding-like elevations―P. aurea DC., P. cappadocica Boiss. & Balansa, P. flabellifolia, P. kotschyana Boiss., P. oliverioides Boiss. & Hausskn. ex Boiss., P. tragium var. pseudotragium (DC.) V.A.Matthews―(fig. 4).
Type 9, ribbed-striate: parallel longitudinal striations with distinct ribbed―P. ibradiensis― (fig. 4G, g).
Cluster analysis of fruit and whole plant morphologyTOP
The results of the cluster analysis of 26 taxa based on fruit morphology as well as whole plant morphology clusters Turkish Pimpinella into 2 groups. Group A contains white-flowered species with the single exception of P. aurea (fig. 5). Group B contains both yellow-flowered and white-flowered species, mostly with glabrous fruits but also a few species with hairy fuits. This group is more heterogeneous than A and includes two differentiated subgroups. Subgroup I contains yellow-flowered species with sparsely hairy fruits whereas subgroup II contains white-flowered species with glabrous fruits, except for P. isaurica V.A.Matthews (fig. 5).
|
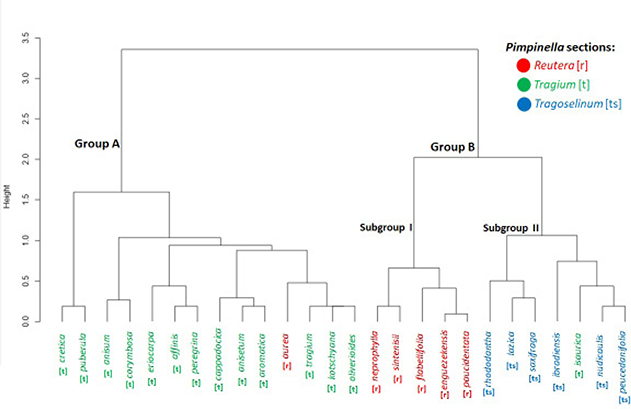 Fig. 5. The cluster dendrogram of the Turkish taxa of Pimpinella L. Fig. 5. The cluster dendrogram of the Turkish taxa of Pimpinella L.
|
|
DISCUSSIONTOP
The first carpological study of Turkish Pimpinella―c. 80% of the taxa―using both SEM and light microscopy has found considerable variation affecting shape and surface. However, variation in shape and surface characters is not correlated and species with similar fruit shapes do not necessarily have similar surface ornamentation. Fruit morphological patterns of variation here analyzed are not fully compatible with classification by Wolff (1927). Phylogenetic relationships are not fully compatible with Wolff’s classification either since the three sections come out as polyphyletic (Magee & al. 2010). However, the combination of fruit and whole plant morphological characters in the cluster analysis provides a partly congruent picture with the classification of Wolff (1927). For instance, our group A includes species from P. sect. Tragium, except for P. aurea, which belongs to P. sect. Reutera. Our subgroup I of group B includes species from P. sect. Reutera whereas subgroup II includes species from P. sect. Tragoselinum except for P. isaurica, which belongs to P. sect. Tragium (fig. 5).
A detailed comparison of Wollf’s classification with the available phylogenetic studies (Tabanca & al. 2005; Magee & al. 2010; Fereidounfar & al. 2016) is hampered by the limited sampling in those studies. However, there is some consistency in the phylogenetic position of the Turkish species across those three studies although with some exceptions―e.gr., P. aurea―. In addition, our micro and macromorpohological study is not fully consistent with the previous anatomical study (Akalın & al. 2016) but a number of associations occurs that is worth commenting, most of which are wholly or partly consistent with the phylogenetic studies. For instance, most species with oblong-ovoid fruits are in the first anatomical group in Akalın & al. (2016). Pimpinella affinis, P. peregrina, and P. eriocarpa all have elliptic fruits and the first two species are in the second anatomical group of Akalın & al. (2016). Our cluster analysis grouped the three species together (fig. 5) and P. peregrina and P. eriocarpa are sister species in the three available phylogenetic studies (Tabanca & al. 2005; Magee & al. 2010; Fereidounfar & al. 2016).
Pimpinella cretica var. cretica and P. puberula share ovoid-globose fruits and other morphological characters (Akalın & al. 2016). Therefore, they come out together in our cluster analysis (fig. 5) and are sister species in Fereidounfar & al. (2016). However, these two species have very different fruit surfaces (fig. 2).
Pimpinella cappadocica, P. anisetum, and P. aromatica share ovoid fruits and are grouped together in the cluster analysis (fig. 5). However, P. anisetum and P. aromatica have rugulose fruit surface whereas that of P. cappadocica is striate. Two of the phylogenetic studies support the closeness of P. cappadocica var. cappadocica and P. anisetum (Tabanca & al. 2005; Magee & al. 2010); the third one does not.
Pimpinella lazica, P. saxifraga, P. enguezekensis, and P. rhodantha all have oblong-ovoid glabrous fruits and came out within group B in the cluster analysis (fig. 5). However, these four species do not share the micromorphological structure of the mericarps since P. lazica and P. saxifraga have rugose-reticulate surface whereas P. enguezekensis and P. rhodantha have it reticulate striate (fig. 3). In contrast, P. saxifraga and P. rhodantha are sister species both in Tabanca & al. (2005) and in Magee & al. (2010); two species that can be distinguished by their flower color as well as their basal and cauline leaves.
Pimpinella nephrophylla, P. sintenisii H.Wolff, and P. paucidentata V.A.Matthews all have oblong fruits, fall within the same cluster―subgroup I of B; fig. 5―and belong to the fourth anatomical group in Akalın & al. (2016), but P. nephrophylla and P. sintenisii differ in their fruit surface (figs. 2, 3). In two of the phylogenetic studies, P. sintenisii and P. paucidentata are closely related (Tabanca & al. 2005; Magee & al. 2010).
Another contrast between morphological and molecular phylogenetic data concerns P. corymbosa and P. kotschyana, which have both ovoid-subglobose fruits but differ in fruit surface and other morphogical characters of the whole plant and yet are closely related in the phylogenetic trees (Tabanca & al. 2005; Magee & al. 2010; Fereidounfar & al. 2016).
The relationships of P. aurea are also controversial. It has a distinct fruit shape (fig. 1) with a striate ornamentation that is similar to P. cappadocica, P. flabellifolia, P. kotschyana, P. oliverioides, and P. tragium var. pseudotragium and falls in the cluster analysis together with P. kotschyana, P. oliverioides, and P. tragium var. pseudotragium. Yet, the phylogenetic position of P. aurea in the two studies in which it was sampled (Tabanca & al. 2005; Magee & al. 2010) differs although in the latter work P. aurea species fell in the same clade as P. cappadocica and P. oliverioides, and the three of them are also closely related to P. kotschyana.
The newly described species P. ibradiensis, which has not been yet included in any phylogenetic study, has been suggested to belong to P. sect. Reutera and to be closely related to P. nephrophylla, P. sintenisii, P. paucidentata, and P. flabellifolia by its authors (Çingilbel & al. 2015). However, our SEM study has found significant differences in micromorphology of fruits (fig. 4) and, in addition, P. ibradiensis can be distinguished from these species by its white petals, serrulate basal leaves, larger fruits, and the presence of bracts and bracteoles. Besides, our cluster analysis placed it together with species of P. sect. Tragoselinum specifically close to P. nudicaulis and P. peucedanifolia.
Our carpological study provides useful previously undetected characters for distinguishing species and, to a lesser degree, for aiding in infrageneric classification of Pimpinella. However, the patterns of variation in fruit micromorphological structures here reported are only partly consistent with our previous anatomical study (Akalın & al. 2016) and with morphological characters of other parts of the plant that are normally used in taxonomy of this genus. This suggests that some of these macro and micromorphological characters may have been acquired independently and thus the information they contain for supporting infrageneric taxonomy of Pimpinella should be ideally confronted to a strongly supported phylogenetic backbone for this genus, which is not yet available.
ACKNOWLEDGEMENTSTOP
We thank to the director of GAZI Herbarium for the fruits of P. oliverioides and Dr. Vahap Eldem for his help with Cluster Analyses. Also we thank to Prof. Gonzalo Nieto Feliner (Real Jardín Botánico) for his suggestions that improved the text. This work was supported by Scientific Research Projects Coordination Unit of Istanbul University, Project numbers: 4821, 53713 and 21369.
REFERENCESTOP
| ○ |
Abebe D. 1992. Systematic studies in the genus Pimpinella L. (Umbelliferae) from tropical Africa. The Journal of Linnean Society Botany 110: 327–372. https://doi.org/10.1111/j.1095-8339.1992.tb00298.x |
| ○ |
Akalın E., Uruşak U. & Kızılarslan Ç. 2013. Fruit anatomy of some Ferulago (Apiaceae) species in Turkey. Turkish Journal of Botany 37 (3): 434–445. |
| ○ |
Akalın E., Yeşil Y. & Akpulat A. 2016. Fruit Anatomy of Turkish Pimpinella Species. Flora 223: 62–73. https://doi.org/10.1016/j.flora.2016.04.004 |
| ○ |
Aksenov E.S. & Tikhomirov V.N. 1972. Klyuch dlya opredeleniya po plodam vidov roda Pimpinella L. flory USSR. Byulleten' Glavnogo Botanicheskogo Sada 85: 35–45. |
| ○ |
Bani B., Ulusoy F., Karakaya M.A. & Koch M.A. 2016a. Taxonomic implications from morphological and anatomical studies in the section Stenodipterafrom the genus Grammosciadium (Apiaceae). PhytoKeys 68: 73–89. https://doi.org/10.3897/phytokeys.68.9089 |
| ○ |
Bani B., Karakaya M.A. & Çeter T. 2016b. Fruit micromorphological characters of the genus Grammosciadium DC. (Apiaceae) in Turkey. Phytotaxa 246 (2): 184–91. https://doi.org/10.11646/phytotaxa.246.3.2 |
| ○ |
Çinbilgel İ., Eren Ö., Duman H. & Gökceoğlu M. 2015. Pimpinella ibradiensis (Apiaceae), an unusual new species from Turkey. Phytotaxa 217 (2):164–172. https://doi.org/10.11646/phytotaxa.217.2.6 |
| ○ |
Engler H.G.A. 1927. Umbelliferae-Apioideae-Blupeurum, Trinia et reliquae Ammineae heteroclitae. In Engler H.G.A. (ed.), Das Pflanzenreich IV 228 (Heft 90). W. Engelmann, Berlin. |
| ○ |
Ertekin A.S. & Kaya Ö.F. 2005. A new record species for the flora of Turkey, Pimpinella nephrophylla Rech. F. & H. Riedl. (Apiaceae). Ot Sistematik Botanik Dergisi 12: 13–18. |
| ○ |
Fereidounfar S., Ghahremaninejad F. & Khajehpiri M. 2016. Phylogeny of the Southwest Asian Pimpinella and related genera based on nuclear and plastid sequences. Genetics and Molecular Research 15 (4): 1–17. https://doi.org/10.4238/gmr15048767 |
| ○ |
Göktürk R.S. 2008. Pimpinella isaurica subsp. sumbuliana (Apiaceae), a new taxon from Turkey. Annales Botanici Fennici 45: 293–295. https://doi.org/10.5735/085.045.0407 |
| ○ |
Hand R. 2011. Euro+Med Plantbase – the information resource for Euro-Mediterranean plant diversity. Website: http://ww2.bgbm.org/EuroPlusMed/ [accessed: 30 April 2018]. |
| ○ |
Khajepiri M., Ghahremaninejad F. & Mozaffarian V. 2010. Fruit anatomy of the Genus Pimpinella L. (Apiaceae) in Iran. Flora 205: 344–356. https://doi.org/10.1016/j.flora.2009.12.030 |
| ○ |
Liu M. & Downie S.R. 2017. The Phylogenetic Significance of Fruit Anatomical and Micromorphological Structures in Chinese
Heracleum Species and Related Taxa (Apiaceae). Systematic Botany 42 (2): 313–325. https://doi.org/10.1600/154823217X695539 |
| ○ |
Lyskov D., Degtjareva G., Samigullin T. & Pimenov M. 2015. Systematic placement of the Turkish endemic genus Ekimia (Apiaceae) based on morphological and molecular data. Turkish Journal of Botany 39: 673–680. https://doi.org/10.3906/bot-1405-111 |
| ○ |
Magee A.R., van Wyk B.E., Tilney P.M. & Downie S.R. 2010. Phylogenetic position of African and Malagasy Pimpinella species and related genera (Apiaceae, Pimpinelleae). Plant Systematics and Evolution 288: 201–211. https://doi.org/10.1007/s00606-010-0325-y |
| ○ |
Matthews V.A. 1972. Pimpinella L. In Davis P.H. (ed.), Flora of Turkey and the East Aegean Islands, vol. 4. University Press, Edinburgh. |
| ○ |
Menemen Y. 2012. Pimpinella L. In Güner A., Aslan S., Ekim T., Vural M. & Babaç M.T. (eds.), Türkiye Bitkileri Listesi (Damarlı Bitkiler). Nezahat Gökyiğit Botanik Bahçesi ve Flora Araştırmaları Derneği Yayını, Istanbul. |
| ○ |
Özcan T. 2004. Analysis of the fruit surfaces in Bupleurum L. (Umbelliferae) with SEM. Plant Systematics and Evolution 247: 61–67. https://doi.org/10.1007/s00606-004-0135-1 |
| ○ |
Pu F.T. & Watson M.F. 2005. Pimpinella L. In Wu Z.Y., Raven P.H. (eds.), Flora of China 14: 93–104. Missouri Botanical Garden Press, Missouri. |
| ○ |
R Development Core Team 2018. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Website: http://www.R-project.org/ [accessed: 12–III–2018]. |
| ○ |
Stearn W.T. 2005. Botanical Latin. David & Charles, Oregon. |
| ○ |
Stevens P.F. 1972. Scaligeria DC. In Davis P.H. (ed.), Flora of Turkey and the East Aegean Islands 4: 333–336. Edinburgh University Press, Edinburgh. |
| ○ |
Tabanca N., Douglas A.W., Bedir E., Dayan F.E., Kirimer N., Baser K.H.C., Aytaç Z., Khan I.A. & Scheffler B.E. 2005. Patterns of essential oil relationships in Pimpinella (Umbelliferae) based on phylogenetic relationships using nuclear and chloroplast sequences. Plant Genetic Resources 3 (2): 149–169. https://doi.org/10.1079/PGR200573 |
| ○ |
Wollf H. 1927. Umbelliferae-Apioideae-Ammineae-Carinae, Ammineae Novemjugatae et Genuinae. In Engler H.G.A. (ed.), Das Pflanzenreich IV 228 (Heft 90): 219–319. W. Engelmann, Berlin. |
| ○ |
Yeşil Y., Yıldırım H., Akalin E., Pirhan A.F. & Altıoğlu Y. 2016. Pimpinella enguezekensis (Apiaceae), a new species from East Anatolia Region (Turkey). Phytotaxa 289 (3): 237–246. https://doi.org/10.11646/phytotaxa.289.3.3 |
| ○ |
Zakharova E.A., Degtjareva G.V. & Pimenov M.G. 2012. Redefined generic limits of Carum (Umbelliferae, Apioideae) and new systematic placement of some of its taxa. Willdenowia 42: 149–168. https://doi.org/10.3372/wi.42.42201 |