Taraxacum sect. Palustria (Compositae) en la península ibérica
Gabriel Mercadal i Corominas 1,* & Antonio Galán de Mera 2
1LAGP-Flora i Vegetació, Herbari de la Universitat de Girona, Facultat de Ciències, campus Montilivi, c/ M. Aurèlia Campmany n.º 69, ES-17003 Girona, España
2Laboratorio de Botánica, Facultad de Farmacia, Universidad CEU San Pablo, P.O. Box 67, ES-28660 Boadilla del Monte, España
*Corresponding author: g.mercadal.corominas@gmail.com, http://orcid.org/0000-0001-6992-8243
2 agalmer@ceu.es, http://orcid.org/0000-0002-1652-5931
| |
Resumen
Mercadal i Corominas G. & Galán de Mera A. 2017. Taraxacum sect. Palustria (Compositae) en la península ibérica. Anales Jard. Bot. Madrid 74 (2): e064. https://doi.org/10.3989/ajbm.2490
Tras la detección de nuevos registros de especies de Taraxacum sect. Palustria (Compositae), se realiza una primera revisión taxonómica de la sección para la península ibérica, donde se reconocen cuatro taxones —T. aginnense, T. ciliare, T. litophyllum y T. raii—, siendo T. aginnense y T. raii novedades para la flora ibérica. De cada una, se aporta su descripción e información sobre su distribución geográfica, preferencias ecológicas, fenología y conservación. Además, se proporciona una clave para su identificación, así como un conjunto de imágenes que muestran los caracteres morfológicos más significativos. El trabajo se complementa incluyendo la delimitación de T. gr. mediterraneum (T. sect. Erythrocarpa), por su similitud morfológica y su afinidad ecológica con sus congéneres ibéricos de T. sect. Palustria.
Palabras Clave. Península ibérica, Taraxacum, Taraxacum sect. Palustria, taxonomía.
|
| |
Abstract
Título en inglés: Taraxacum sect. Palustria (Compositae) in the Iberian Peninsula.
After detecting new records for Taraxacum sect. Palustria (Compositae), a first taxonomical revision of this section is provided for the Iberian Peninsula. Four taxa, T. aginnense, T. ciliare, T. litophyllum and T. raii, are recognized, of which T. aginnense and T. raii are new for the Iberian flora. From each one of them, its morphological description, geographical, ecological and phenological data, and conservation status are presented. In addition, an identification key, as well as several images showing the main morphological characters are given. This work also includes comments on T. mediterraneum group (T. sect. Erythrocarpa) because its morphological similitude and ecological affinities with Iberian plants of T. sect. Palustria.
Keywords. Iberian Peninsula, Taraxacum, Taraxacum sect. Palustria, Taxonomy.
|
INTRODUCCIÓNTOP
Taraxacum sect. Palustria (H.Lind.) Dahlst. comprende en la actualidad más de 140 especies (Kirschner & al. 2017) de suelos húmedos cuyas características
comunes son: brácteas externas del involucro ovadas u ovado-lanceoladas, adpresas, erecto-patentes o ligeramente recurvadas,
de tonalidades rosadas o purpúreas, con un margen escarioso bastante ancho, los aquenios de un verde amarillento a verde oliva,
incluso pardos, y las hojas dentadas o con lóbulos simples (Dahlstedt 1921; Kirschner & Štěpánek 1998; Galán de Mera 2017).
Algunos autores, como Wittzell (1999) sugieren que T. sect. Palustria, junto con T. sect. Alpina G.E.Haglund, T. sect. Arctica Dahlst., T. sect. Erythrocarpa Hand.-Mazz. y T. sect. Scariosa Hand.-Mazz., forman un grupo de especies predominantemente diploides y sexuales, precursoras de otras poliploides y agamospérmicas,
como las de T. sect.Taraxacum. Sin embargo, en T. sect. Palustria se han encontrado especies con el polen irregular y reproducción agamospérmica que presentan una morfología muy variada (Uhlemann 2001; Vašut 2003), pues, dentro del género, son raras las secciones puramente sexuales (Kirschner & al. 2015). En otros casos, se han detectado especímenes hibridógenos, muy próximos por morfología y genoma de cloroplastos a otras
secciones, como T. sect. Erythrocarpa o T. sect. Taraxacum (Kirschner & al. 2003; Záveský & al. 2007), aunque los caracteres antedichos de T. sect. Palustria son más o menos homogéneos.
El centro de diversificación de T. sect. Palustria se encuentra en Europa central y oriental, quedando la península ibérica muy al margen de esta área (Kirschner & Štĕpánek 1998). En la Península solo se habían confirmado dos taxones, T. ciliare Soest y T. litophyllum De Langhe & Soest. El primero se consideraba la única especie peninsular de la sección, con las localidades de Grazalema
—Cádiz—, Sierra Nevada —Granada— y San Juan de la Peña —Huesca—; T. litophyllum, con solo la localidad típica de Aigües Tortes —Lleida— (De Langhe & Soest 1972), no era considerada en la monografía de Kirschner & Štĕpánek (1998).
El presente estudio es la primera revisión de Taraxacum sect. Palustria para la península ibérica, y responde a una ampliación de la información sobre el género Taraxacum F.H.Wigg. en Flora iberica (Galán de Mera 2017). Se reconocen cuatro taxones: T. aginnense Hofstra, T. ciliare, T. litophyllum y T. raii (Gouan) S.F.Gray. Además, se incluyen también poblaciones de Taraxacum gr. mediterraneum —T. sect. Erythrocarpa—, que presentan muchas similitudes morfológicas y ecológicas con T. aginnense.
MATERIAL Y MÉTODOSTOP
El estudio biométrico en el que se fundamenta esta contribución ha sido llevado a cabo con material recolectado por los autores,
y con el conservado en los herbarios BR, HGI, JACA, L, LG, MA, MAF, PRA, USP y VAL —acrónimos recogidos en Thiers (2017)—. En el caso de T. aginnense y T. raii, el estudio se ha basado exclusivamente en material fresco que posteriormente se ha conservado en HGI. A partir de los datos
obtenidos, se aporta una clave dicotómica, una descripción detallada de cada taxón —que se acompaña de imágenes que muestran
su variabilidad morfológica— y un mapa de distribución geográfica en Europa y la península ibérica —precisión de recuadro
UTM de 10 km de lado—. Los mapas se han construido a partir de datos de campo propios, testimonios de herbario y referencias
bibliográficas. Los pliegos y las referencias usadas se detallan en la descripción de cada taxón. La información sobre el
número de cromosomas procede de la base de datos de Watanabe (2017).
Para conocer el comportamiento ecológico de las plantas recolectadas, se han estudiado distintas muestras de suelos recogidas
durante la primavera —de marzo a mayo— en prados donde habitan T. aginnense, T. ciliare, T. raii y T. gr. mediterraneum del NE de España y el SE de Francia, analizándose los factores que más influyen sobre la vegetación: la textura —granulometría
por sedimentación discontinua: método de la pipeta de Robinson—, el contenido de humedad de campo —método gravimétrico—, el
pH 1:2,5 en H2O, la conductividad eléctrica 1:5 a 25 ºC, la materia orgánica —método de oxidación por vía fría— y el contenido de carbonato
cálcico equivalente —método del calcímetro de Bernard—. Todos los análisis se han realizado según los protocolos indicados
por Porta & al. (1986, 2003). Los resultados se sintetizan en las tablas 1–4. Finalmente, se aporta una tabla con las principales características morfológicas y ecológicas distintivas de cada taxón
(apéndice 1).
Tabla 1. Características edáficas de los prados donde habita Taraxacum aginnense Hofstra. [ISSS, International Society of Soil Science.]
| Valores |
pH 1:2,5 H2O
|
CE 1:5 a 25ºC (µS/cm) |
Carbonatos (%) |
Materia orgánica (%) |
Humedad de campo (%) |
Clase textural [ISSS] (número de muestras) |
| Mínimo |
7,1 |
124,8 |
0,0 |
2,2 |
20,9 |
arcillosa arenosa (1) |
| Máximo |
8,4 |
421,0 |
13,4 |
4,8 |
33,0 |
arcillosa fina (2) |
| Media |
7,4 |
303,1 |
4,4 |
3,6 |
27,9 |
arcillosa gruesa (1) |
| Desviación estándar |
1,2 |
106,2 |
6,1 |
1,2 |
4,6 |
— |
| Número de muestras |
6 |
6 |
4 |
4 |
6 |
— |
RESULTADOSTOP
Clave de especies de T. sect. Palustria de la península ibérica, incluyendo T. gr. mediterraneum (T. sect. Erythrocarpa)TOP
| 1. |
Brácteas externas erectas, patentes o ligeramente recurvadas, de lanceoladas a ovado-lanceoladas, habitualmente con una relación entre el largo y el ancho ≧ (1,9)2,2; longitud del cono de los aquenios 0,4–0,8 mm; polen de tamaño irregular |
5. T. gr. mediterraneum |
| – |
Brácteas externas aplicadas o erectas (raramente patentes), de ovadas a ovado-lanceoladas, habitualmente con una relación entre el largo y el ancho ≦ 2,2(2,5); longitud del cono de los aquenios (0,3)0,4–1,1; polen nulo o de tamaño regular o irregular |
2 |
| 2. |
Flores sin polen; vilano discoloro —blanco, verdoso en la base—; pecíolo alado; corola de color amarillo azafranado; hojas poco variables, enteras o ligeramente dentadas |
3. T. litophyllum |
| – |
Flores generalmente con polen; vilano concoloro —blanco—; pecíolo habitualmente áptero; corola amarilla; hojas muy variables, dentadas o de pinnatífidas a pinnatisectas |
3 |
| 3. |
Granos de polen —véanse a 600×— de tamaño casi regular —< 10% de los granos son desiguales—; aquenios con conos (0,4)0,5–0,8(0,9) mm de longitud |
4. T. raii |
| – |
Granos de polen —véanse a 600×— de tamaño irregular —≧ 20% de los granos son desiguales— o, raramente, ausentes; aquenios
con conos de (0,3)0,4–1,1 mm de longitud |
4 |
| 4. |
Aquenios con cono de (0,3)0,4–0,6(0,7) mm de longitud; granos polen de distintos tamaños —a veces, la diferencia se observa solo a partir de 600×—; brácteas externas adpresas, erectas, raramente patentes, ovado-lanceoladas, a veces ovadas, con una relación entre el largo y el ancho (1,3)1,7–2,2(2,5) mm; hojas habitualmente de pinnatífidas a pinnatisectas —rara vez dentadas—, ordinariamente con el segmento terminal con una relación entre el largo y el ancho 0,7–1,5 |
1. T. aginnense |
| – |
Aquenios con cono de (0,6)0,8–1,1 mm de longitud; granos de polen de distintos tamaños —la diferencia se suele observar a partir de 60×—; brácteas externas fuertemente adpresas o ligeramente erectas, ovadas, a veces ovado-lanceoladas, con una relación entre el largo y el ancho 1,1–2,2(2,3) mm; hojas espatulado-lanceoladas, dentadas o de pinnatífidas a pinnatisectas, si hay segmento terminal, habitualmente con una relación entre el largo y el ancho de (0,6)1,0–2,2 |
2. T. ciliare |
Tratamiento taxonómicoTOP
Taraxacum sect. Palustria (H.Lind.) Dahlst.
1. T. aginnense Hofstra, Bull. Soc. Bot. Fr., Lett. Bot. 132: 130 (1985). Tipo: [Francia], Aquitaine, Lot-et-Garonne, NE Grayssas y SW Pissot,
1-IV-1981, J. Hofstra s.n. (holo-: L 0214958!). Fig. 1.
|
 Fig. 1. Taraxacum aginnense Hofstra: a, hábito; b, capítulo con la brácteas externas de erectas a patentes; c, detalle de la hoja pinnatisecta; d, polen de tamaño irregular junto a los tejidos de una antera; e-g, variabilidad de las brácteas externas; h-j, variabilidad del cuerpo y cono de los aquenios [a, b, HGI 23525; c, h, i, HGI 23418; d, g, HGI 23526; e, f, j, HGI 23445]. Fig. 1. Taraxacum aginnense Hofstra: a, hábito; b, capítulo con la brácteas externas de erectas a patentes; c, detalle de la hoja pinnatisecta; d, polen de tamaño irregular junto a los tejidos de una antera; e-g, variabilidad de las brácteas externas; h-j, variabilidad del cuerpo y cono de los aquenios [a, b, HGI 23525; c, h, i, HGI 23418; d, g, HGI 23526; e, f, j, HGI 23445].
|
|
Hierba hasta de 50 cm. Tallos erectos, más cortos o más largos que las hojas, sin hojas bracteiformes, pubescentes, con indumento
aracnoideo o lampiños al madurar, verdes, pardos o purpúreos. Hojas 6–30,0 × 1,0–4,5 cm, pecioladas, espatulado-lanceoladas,
de pinnatífidas a pinnatisectas, raramente dentadas, suaves, verdes, sin manchas purpúreas, solo a veces ligeramente entre
los lóbulos; nervio medio sin estrías longitudinales, verde o purpúreo; lóbulos laterales (0)2–5(7) por hemilimbo, de 5,0–28,0
× 0,4–21 mm —relación entre el largo y el ancho 1,0–2,3—, linear-triangulares, triangulares, deltoideos, indivisos, con el
margen distal entero, recto o sigmoideo y el proximal entero, recto o ligeramente sigmoideo, con los dientes 1–2 mm, triangulares,
con o sin dientes entre los lóbulos siendo más comunes hacia la parte basal, los proximales se continúan en el pecíolo con
algún diente triangular; lóbulo terminal (1,0)8,0–38,0 × (1,0)8,0–32,0 mm —relación entre el largo y el ancho 0,7–1,5—, obtuso,
frecuentemente triangular, generalmente con dientes distantes y los márgenes de la base generalmente asimétricos; pecíolo
de verde a purpúreo, estriado longitudinalmente, áptero o más raramente alado, a menudo con dientes. Capítulo 27–38 mm de
diámetro en la antesis. Involucro 12–25 × 4–15 mm; brácteas externas 4,5–8,4 × 2,2–4,8 mm, ovado-lanceoladas, a veces ovadas
—relación entre el largo y el ancho de (1,3)1,7–2,2(2,5)—, adpresas, erectas o raramente patentes, especialmente en la madurez,
con margen escarioso de 0,1–0,4 mm de anchura, de un verde obscuro, frecuentemente teñidas de púrpura hacia los márgenes y
el ápice, cilioladas, sin cuerno o callo subapical. Flores hermafroditas. Corola hasta de 20 mm de longitud, de amarilla pálida
a amarilla, las más externas del capítulo con una banda ancha rojiza en el dorso. Anteras amarillas, con granos de polen de
distintos tamaños. Ramas estilares de un amarillo verdoso, de pardo a castaño obscuro cuando secas. Aquenios de color pardo
claro; cuerpo (2,0)3,0–4,0 mm de longitud, con escuámulas erosas bi o tricuspidadas, y espículos cortos hacia el ápice, el
resto liso, con las costillas ± anchas; cono (0,3)0,4–0,6(0,7) mm, de cónico a subcilíndrico; pico 5,0–8,3(10,0) mm de longitud,
blanquecino. Vilano 4,5–7,0(8,2) mm de longitud, concoloro, blanco.
Hábitat.—Habita en prados y herbazales de mesohigrófilos a higrófilos, principalmente en prados de guadaña temporalmente inundados
—Trifolio-Hordeetalia Horvatic 1963—, herbazales de Oenanthe fistulosa —Ranunculo-Oenanthion de Foucault in de Foucault & Catteau 2012— y pastos nitrificados —Trifolio-Cynodontion Br.-Bl. & O.Bolòs 1958—. En Francia, Hofstra (1985) indica esta planta en el Agropyro-Rumicion Nordhagen 1940. Resiste el pastoreo temporal. Los suelos son preferentemente arcillosos y normalmente paraturbosos, de reacción
neutra a básica y siempre temporalmente inundados (tabla 1). En la península ibérica se ha recolectado entre 0–200 m de altitud, y en el departamento de Vaucluse —Provenza—, Girerd & Roux (2011) la han observado hasta los 350 m.
Fenología.—De marzo a mayo.
Distribución.—Europa occidental: Bélgica (Lambinon & Verloove 2012; Kirschner & Štĕpánek 1998), Francia (Hofstra 1985, 1990; Kirschner & Štĕpánek 1998; Girerd & Roux 2011; Tison & Foucault 2014; Tison & al. 2014; Molina 2015; Noble & al. 2016; Tison 2017) y España. En la península ibérica se ha herborizado hasta ahora en el noreste en Cataluña, en las comarcas gerundenses del
Alt Empordà, Baix Empordà, Pla de l’Estany y la Selva, y en la barcelonesa del Maresme. Más al norte, también se ha recolectado
en el Rosselló —Francia— (fig. 2).
Conservación.—Kirschner & Štĕpánek (1998) consideraron este taxón como endémico —EN— de Bélgica y Francia, pero T. aginnense penetra como mínimo hasta el noroeste de España. Tanto Molina (2015) como Noble & al. (2016) lo consideran sin datos suficientes —DD— en las regiones Languedoc-Roussillon y Provence-Alpes-Côte d’Azur respectivamente.
En Cataluña esta planta aún se ha colectado poco y no se dispone de bastante información para asignarle un estatus IUCN, así
pues también se considera sin datos suficientes —DD.
Observaciones.—Como han indicado previamente Kirschner & Štĕpánek (1998), Tison & al. (2014) y Tison & Foucault (2014), T. aginnense corresponde a una especie marginal dentro de T. sect. Palustria, tanto por sus hojas extremadamente divididas, de pinnatipartidas a pinnatisectas, que recuerdan a T. sect. Erythrosperma (H.Lindb.) Dahlst. y T. sect. Erythrocarpa, como por los conos de los aquenios que son muy pequeños, generalmente < 0,6 mm de longitud. Además, de las cuatro especies
tratadas dentro de T. sect. Palustria, esta es la que tolera suelos más nitrificados y alterados, lo que coincide con su polen irregular y el comportamiento reproductivo
agamospérmico (Uhlemann 2001). Es un taxón cercano morfológicamente a T. gr. mediterraneum (T. sect. Erythrocarpa) del cual resulta a menudo difícil de distinguir. Taraxacum aginnense tiende a presentar las brácteas externas ovado-lanceoladas, a veces ovadas, con una relación entre el largo y el ancho habitualmente
≦ 2,2, adpresas o erectas —raramente patentes—; mientras que en T. gr. mediterraneum tienden a ser de lanceoladas a ovado-lanceoladas, con una relación entre el largo y el ancho habitualmente ≧ 2,2, erectas,
patentes o ligeramente recurvadas. Respecto al polen, el ≧ 20% de los granos son desiguales, pero a veces este carácter solo
es distinguible a partir de los 600×.
Material estudiado.—ESPAÑA. Girona: Banyoles, vora el mas Arbeix, 31TDG8064, 174 m, 26-III-2017, G. Mercadal (HGI 23509); Bellcaire d’Empordà, a Sobrestany, 31TEG1059, 3 m, 12-IV-2016, G. Mercadal (HGI 23446); Castelló d’Empúries, a les closes de la Paborderia, 31TEG0679, 4 m, 12-IV-2016, G. Mercadal (HGI 23427); Pau, a les closes de Mornau, 31TEG0882, 1 m, 5-V-2016, G. Mercadal (HGI 23441); Sils, als Tres Ponts, 31TDG8028, 66 m, 22-III-2016, G. Mercadal (HGI 23418); Sils, als Tres Ponts, 31TDG7926, 66 m, 15-IV-2017, G. Mercadal (HGI 23526); Sils, als Tres Ponts, 31TDG8028, 67 m, 15-IV-2017, G. Mercadal (HGI 23525).
FRANCIA. Pyrénées-Orientales: Tuïr, a la Prada, 31TDH8120, 85 m, 7-IV-2015, G. Mercadal (HGI 23445).
2. T. ciliare Soest, Acta Bot. Neerl. 14: 25 (1965). Tipo: France, Vosgos, Gerbamont, V-1887, D. Pierrat s.n. (holo-: L 0002403!). Fig. 3.
|
 Fig. 3. Taraxacum ciliare Soest: a, hábito; b, capítulo con las brácteas externas adpresas; c-f, variabilidad de las brácteas externas; g-i, variabilidad del cuerpo y cono de los aquenios; j, polen de tamaño irregular; k-ñ, variabilidad de las hojas [a, b, f, h, k, HGI 23437; c, i, HGI 23504; d, HGI 18983; e, m, HGI 23506; g, HGI 18984; j, n,
HGI 23435; l, HGI 23436; ñ, HGI 23505]. Fig. 3. Taraxacum ciliare Soest: a, hábito; b, capítulo con las brácteas externas adpresas; c-f, variabilidad de las brácteas externas; g-i, variabilidad del cuerpo y cono de los aquenios; j, polen de tamaño irregular; k-ñ, variabilidad de las hojas [a, b, f, h, k, HGI 23437; c, i, HGI 23504; d, HGI 18983; e, m, HGI 23506; g, HGI 18984; j, n,
HGI 23435; l, HGI 23436; ñ, HGI 23505].
|
|
T. hoedicense Soest, Acta Bot. Neerl. 14: 32 (1965), nom. inval. —McNeill & al. (2012): art. 9.1, 37.1.
T. sarniense A.J.Richards, Watsonia 15: 93 (1984)
Hierba hasta de 50 cm. Tallos erectos, más cortos o más largos que las hojas, sin hojas bracteiformes, pubescentes, con indumento
aracnoideo, lampiños al madurar, quedando en ocasiones algo de pelosidad bajo el capítulo, verdes, pardos o purpúreos. Hojas
1,2–13,5 × 0,3–1,9 cm, pecioladas, espatulado-lanceoladas, dentadas o de pinnatífidas a pinnatisectas, suaves, verdes, sin
manchas purpúreas; nervio medio sin estrías longitudinales, verde o purpureo; lóbulos laterales 3–5(7) por hemilimbo, de (1,0)3,1–11,9
× 1,6–12,4 mm —relación entre el largo y el ancho 0,5–1,8—, de triangulares a deltoideos, indivisos, con el margen distal
entero, recto o sigmoideo y el proximal entero, recto o ligeramente sigmoideo, con los dientes 1–2 mm, triangulares, sin dientes
entre los lóbulos, los proximales se continúan en el pecíolo con algún diente triangular; lóbulo terminal 11,6–41,4 × (2,7)5,0–15,6
mm —a veces falta; relación entre el largo y el ancho (0,6)1,0–2,2(4,4)—, agudo, frecuentemente lingüiforme, generalmente
con dientes distantes y los márgenes de la base simétricos; pecíolo de verde a purpureo, estriado longitudinalmente, áptero,
con o sin pequeños dientes. Capítulo de 25–35(40) mm de diámetro en la antesis. Involucro 9,9–16,9 × 4,3–8,4 mm; brácteas
externas (2,8)4,3–8,0(9,1) × (1,6)2,4–5,3 mm —relación entre el largo y el ancho 1,1–2,2(2,3) mm—, de ovadas a ovado-lanceoladas,
adpresas o ligeramente erectas, con margen escarioso de 0,1–1,1 mm de anchura, verde o verde obscuro, frecuentemente teñidas
de púrpura hacia los márgenes y el ápice, cilioladas, sin cuerno o callo subapical. Flores hermafroditas. Corolas 6,7–18 mm
de longitud, de color amarillo intenso, las más externas del capítulo con una banda ancha rojiza en el dorso. Anteras amarillas,
con granos de polen de distintos tamaños o, muy raramente, ausentes. Ramas estilares de un verde amarillento. Aquenios de
un verde amarillento a verde oliva; cuerpo 2,7–4,0 mm de longitud, con escuámulas erosas bi o tricuspidadas, o con espículos
cortos hacia el ápice, el resto liso, con las costillas ± anchas; cono (0,6)0,8–1,1 mm de longitud, de subcilíndrico a cilíndrico,
pico 4,5–9,5 mm de longitud, de un verde amarillento. Vilano 3,8–7,2 mm de longitud, concoloro, blanco.
Hábitat.—Crece principalmente en prados subsalinos litorales —Juncion maritimi Br.-Bl. ex Horvatic 1934, Trifolion maritimi Br.-Bl. in Br.-Bl., Roussine & Négre 1952—, herbazales higrófilos —Ranunculo-Oenanthion, Oenanthion fistulosae de Foucault 2008 em. de Foucault in de Foucault & Catteau 2012— y prados de siega temporalmente inundados —Trifolio-Hordeetalia—. También se puede encontrar esporádicamente en fresnedas inmaduras —Carici-Fraxinion oxycarpae Pedrotti ex Pedrotti & al. 2014— que corresponden a antiguos prados abandonados (Mercadal & al. 2003, 2008; Mercadal & Vilar 2013). En el interior de España se ha herborizado en gramadales subsalinos inundables —Trifolio-Cynodontion—, mientras que en el sur de Francia Hofstra (1985) la recolectó en herbazales higrófilos nitrificados del Agropyro-Rumicion. Los suelos son preferentemente de arcillosos a arcillo-arenosos, paraturbosos, de reacción indiferente, pero frecuentemente
básicos, incluso subsalinos, y habitualmente inundados hasta entrada la primavera (tabla 2). Altitudinalmente se distribuye desde el nivel del mar al piso montano, de 0–1900 m.
Tabla 2. Características edáficas de los prados donde habita Taraxacum ciliare Soest. [ISSS, International Society of Soil Science.]
| Valores |
pH 1:2,5 H2O
|
CE 1:5 a 25ºC (µS/cm) |
Carbonatos (%) |
Materia orgánica (%) |
Humedad de campo (%) |
Clase textural [ISSS] (número de muestras) |
| Mínimo |
5,9 |
70,0 |
0,0 |
2,7 |
21,7 |
arcillosa arenosa (2) |
| Máximo |
8,2 |
1497,0 |
9,0 |
4,9 |
60,0 |
arcillosa fina (3) |
| Media |
7,1 |
190,7 |
1,3 |
3,3 |
30,5 |
arcillosa gruesa (2) |
| Desviación estándar |
0,8 |
411,4 |
3,4 |
0,8 |
10,4 |
— |
| Número de muestras |
12 |
12 |
7 |
7 |
12 |
— |
Fenología.—De marzo a junio.
Distribución.—Europa occidental: Holanda (Soest 1965), Islas del Canal de la Mancha (Richards & Haworth 1984; Sell & Murrell 2006), Francia (Hofstra 1985; Kirschner & Štĕpánek 1998; Girerd & Roux 2011; Lambinon & Verloove 2012; Tison & Foucault 2014; Molina 2015; Noble & al. 2016; Tison 2017) y España (Kirschner & Štĕpánek 1998; Mercadal & Vilar 2013; Galán de Mera 2017), norte de Italia (Kirschner & Štĕpánek 1998; Aquaro & al. 2008; Soldano 2010) y sureste de Suiza (Kirschner & Štĕpánek 1998). Muy cerca del núcleo poblacional de la región Rhône-Alpes de Francia se conocen formas aberrantes que también deben corresponder
a T. gr. ciliare. En la península ibérica se encuentra disperso, pero es más frecuentemente en la mitad oriental, desde las comarcas gerundenses
del Alt Empordà y de la Selva, Pirineos Centrales —Huesca— y País Vasco —Araba-Álava—, al Sistema Ibérico —Soria, Cuenca,
Guadalajara e interior de Valencia— y sierras de Castellón; por el sur, se distribuye por los humedales montanos desde Granada
a Cádiz; y en la meseta norte, se encuentra en las proximidades de las lagunas de Tierra de Campos —Valladolid y Zamora—.
Su distribución en la Península es en buena parte paralela a la de los humedales subsalinos interiores. Más al norte de España,
también se ha colectado en el Rosselló y en el Conflent (fig. 4).
Conservación.—Kirschner & Štĕpánek (1998) consideran este taxón como vulnerable —VU— a escala europea. Hawksworth (2002), incluyendo citas de T. sarniense, lo da por probablemente extinto —EX— en las Islas del Canal de la Mancha —Guersney y Jersey—. En el norte de Francia, incluyendo
citas de T. hoedicense, se considera en fuerte regresión (Lambinon & Verloove 2012; Tison & Foucault 2014), y en el sur, Molina (2015) y Noble & al. (2016) no le han asignado ningún estatus de conservación IUCN por poseer pocos datos —DD— de las regiones de Languedoc-Roussillon
y Provence-Alpes-Côte d’Azur respectivamente. Aun así, probablemente el estatus más idóneo para toda su área de distribución
es el de casi amenazado —NT—. En la península ibérica, donde existen varios núcleos repartidos por buena parte del territorio,
lo consideramos también como casi amenazado —NT—, ya que tiene una distribución amplia; pero los distintos núcleos poblacionales
tienden a desaparecer por el saneamiento y la urbanización de las zonas húmedas donde se desarrolla. Además, cabe destacar
que debido a la complejidad del género, este taxón, como la mayoría de sus congéneres de T. sect. Palustria, se ha debido confundir generalmente con los de otras secciones, especialmente de T. sect. Taraxacum, y está subestimado.
Observaciones.—Bajo el binomen T. ciliare englobamos a un conjunto de poblaciones que presentan unas características bien distinguibles del resto de congéneres de
la misma T. sect. Palustria en la península ibérica: cono alargado —0,6–1,1 mm—, polen de tamaño claramente irregular —≧ 20% de los granos son desiguales—
al observarlo a partir de 60×, y brácteas externas de ovadas a ovado-lanceoladas, con una relación entre el largo y el ancho
de 1,1–2,2(2,3). La mayoría de los ejemplares ibéricos son muy parecidos a la planta típica de la regiones atlántica y subatlántica
de Francia —locus classicus de T. hoedicense— e Islas del Canal de la Mancha —locus classicus de T. sarniense—: brácteas cortas y anchas, conos de subcilíndricos a cilíndricos > 0,8 mm, hojas de dentadas a pinnatífidas, estas últimas
con un largo segmento apical con forma de obús —ver iconografía en Kirschner & Štĕpánek (1998: 28)—. Pero algunos otros ejemplares presentan los conos más cortos, subcónicos, de tan solo 0,6 mm de longitud, y las brácteas
externas a veces son más bien ovado-lanceoladas y erectas, por lo cual Kirschner & Štĕpánek (1998) los considerarían como formas aberrantes o atípicas. La forma típica de T. ciliare se distribuye de manera dispersa por el continente, siendo más frecuente en Holanda, Bélgica y el NO de Francia, rodeando
al locus classicus de T. ciliare, en tanto que las formas atípicas se distribuirían principalmente en localidades más meridionales de Europa occidental, como
NE de la península ibérica y S de Francia, Suiza y el norte de Italia. Todas las poblaciones estudiadas presentan el polen
de tamaño irregular, lo que indica un comportamiento reproductivo agamospérmico. Uribe-Echebarría (1999) citó T. palustre (Lyons) Symons, Syn. Pl. Ins. Brit.: 172 (1798) [Leontodon palustre Lyons, Fasc. Pl. Cantabr.: 48 (1763), basón.; neotipo designado por Kirschner & Štĕpánek (1998: 31): OXF] en el País Vasco; sin embargo, el material que hemos visto de este territorio no presenta las características
que Kirschner & Štĕpánek (1998) indican para esta especie —brácteas externas del involucro con el margen escarioso de 0,1–0,2 mm, las flores sin polen y
el cuerpo de los aquenios de 4,2–4,7 mm—, sino las de T. ciliare. También, las referencias a T. lividum (Waldst. & Kit.) Peterm., Deutschl. Fl.: 337 (1844) [Leontodon lividus Waldst. & Kit., Descr. Icon. Pl. Hung.: 120 (1802), basón.], y a T. paludosum (Scop.) Crep., Man. Fl. Belgique ed. 2: 231 (1866) [Hedypnois paludosa Scop., Fl. Carniol. ed. 2, 2: 100, tab. 48 (1772), basón.; lectotipo designado por Kirschner & Štĕpánek (1998: 185): Scopoli (1772: tab. 48), icon] de Richards & Sell (1984) en España, o a T. officinale var. lividum W.D.J.Koch, Syn. Fl. Germ. Helv.: 428 (1837) y T. salinum sensu Willk. (Willkomm & Lange 1865) en Aragón, Llobregat —Barcelona— y Trasierra —Córdoba—, seguramente deban llevarse a T. ciliare.
Material estudiado.—ESPAÑA. Barcelona: Tordera, al pla d’Anyells, al prat de can Gener, 31TDG7618, 32 m, 19-III-2017, G. Mercadal (HGI 23506). Cádiz: Grazalema, Sierra de Zafalgar, 30STF8374, 1000–1100 m, 1-V-1994, O. Šída (PRA 88). Castellón/Castelló: Cedramán, 30TYK2040, 700 m, 5-IV-1987, A. Nebot (VAL 62107); Puebla de Benifasar, 31TBF0609, 700 m, 5-V-1990, G. Mateo 1990 (VAL 67228). Cuenca: Garaballa, macizo del Mazmorra, 30SXK4050, 1200 m, 3-IV-1997, Mateo y Pascual 12973 (VAL 105933). Girona: Agullana, al prat d’Eina, 31TDG8692, 217 m, 9-IV-2011, G. Mercadal (HGI 22102); Caldes de Malavella, a ca n’Oms, 31TDG8832, 107 m, 25-III-2001, G. Mercadal (HGI 18984); Caldes de Malavella, als prats de Sant Sebastià, 31TDG8230, 89 m, G. Mercadal (HGI 23527); Cantallops, 31TDG9394, 165 m, 22-III-2003, G. Mercadal (HGI 18983); Cantallops, a la Verneda, 31TDG9394, 160 m, 11-IV-2003; G. Mercadal (HGI 18995). Granada: Sierra Nevada, vía pública GR 420 Granada-Pico Veleta, Fuente de Don Manuel, 30SVG 5612, 1900 m, 23-VI-1993,
Z. Skála (PRA 86, PRA 87). Guadalajara: Orea, hacia Griegos, 30TXK0982, 1650 m, 13-VI-1988, G. Mateo 382 (VAL 58962). Huesca: Aisa, manantial junto al río Estarrún, cerca del refugio de Igüer, 30TXN9735, 1580 m, 18-VI-1986, P. Montserrat y J.L. Remon (JACA 415986); Aisa, Igüer, El Tremedal, 30TXN9736, 1620 m, 18-VI-1986, P. Montserrat y J.L. Remon (JACA 417986); Monasterio de San Juan de la Peña, 30TXN9109, 1250 m, 21-V-1972, P. Auquier 1791, E. Favaux y J. Lambinon (JACA s.n., LG); Yebra de Basa, Santa Osoria, 30TYN2311, 1550 m, 23-IV-2003, P. Montserrat (JACA R274124). Soria: Beratón, nacedero del río Iruela, 30TXM0020, 1550–1600 m, 10-V-2000, I. Blinova, D. Gómez y P.M. Uribe-Echevarría (JACA 15000); Cabrejas del Campo, 30TWM6315, 1000 m, 30-IV-1975, A. Segura Zubizarreta (MA 478901). Valencia/València: Arroyo Cerezo, 30TXK3343, 1380 m, 1-V-1988, G. Mateo (VAL 57767). Valladolid: Encinas de Esgueva, 30TVM0824, 833 m, 5-IV-1983, J.L. Fernández Alonso (MA 294913). Álava/Araba: Maestu, Korres, barranco Monasterio, 30TWN4628, 650–700 m, 18-IV-2000, D. Gómez (JACA 140700); Maestu, Apellaniz, Los Arrancados, 30TWN4030, 770–800 m, 18-IV-2000, D. Gómez, P. Uribe y C. Aseguinolaza (JACA 140500). Zamora: Lagunas de Villafáfila, 30TTM8438, 686 m, 25-III-2017, E. Linares y A. Galán 4616 (USP).
FRANCIA. Pyrénées-Orientales: Montalban, al prat del Camp del Roc, 31TDH6427, 441 m, 31-III-2017, G. Mercadal (HGI 23504); Rodès, vora el mas Batlle, al prat de la Font, 31TDH6125, 492 m, 31-III-2017, G. Mercadal (HGI 23505); Salses, a les closes del Garrigot, 31TDH9640, 0 m, 7-IV-2016, G. Mercadal (HGI 23438); Salses, a les closes del Garrigot, 31TDH9640, 0 m, 7-IV-2016, G. Mercadal (HGI 23436); Sant Nazari, als prats de l’estany, 31TDH9923, 1 m, 31-III-2016, G. Mercadal (HGI 23435); Sant Nazari, a l’antic estany, 31TDH9924, 1 m, 31-III-2016, G. Mercadal (HGI 23437).
3. T. litophyllum De Langhe & Soest, Bull. Soc. Roy. Bot. Belgique 105: 115, fig. 1 (1972). T. palustre subsp. litophyllum (De Langhe & Soest) O. Bolòs & al., Fl. Manual Països Catalans: 1215 (1990). Tipo: España, [Lleida], Parque Nacional de Aigües
Tortes, 1200 m, 8-VII-1970, J. De [Langhe] 257 (holo-: BR 970947!). Fig. 5.
|
 Fig. 5. Taraxacum litophyllum De Langhe & Soest: a, hábito; b, c, ampliación de los capítulos, donde se observan las brácteas externas y los vilanos discoloros [BR 970947; fotografía por
cortesía del jardín Botánico de Meise (Agentschap Plantentuin Meise)]. Fig. 5. Taraxacum litophyllum De Langhe & Soest: a, hábito; b, c, ampliación de los capítulos, donde se observan las brácteas externas y los vilanos discoloros [BR 970947; fotografía por
cortesía del jardín Botánico de Meise (Agentschap Plantentuin Meise)].
|
|
Hierba hasta de 35 cm. Tallos erectos, más largos que las hojas, lampiños, de verdes a pardos o ligeramente purpúreos. Hojas
4–11 × 1–1,5 cm, pecioladas, espatulado-lanceoladas, enteras o con pequeños dientes distantes, suaves, verdes, sin manchas
purpúreas; nervio medio sin estrías longitudinales, de pardo a purpúreo; pecíolo de pardo a purpúreo, sin estrías longitudinales,
alado, sin dientes. Capítulo de c. 30 mm de diámetro en la antesis. Involucro 13,4–14,2 × 6,2–6,5 mm; brácteas externas 4,7–7,5
× 1,1–2,3 mm, ovado-lanceoladas —relación entre el largo y el ancho de 3,2–4,3—, adpresas, con margen escarioso de 0,1–0,4
mm de anchura, de color verde oliva, a veces cilioladas, sin cuerno o callo subapical. Flores hermafroditas. Corolas c. 20
mm de longitud, de color amarillo azafranado, las más externas del capítulo con una banda ancha cano-purpúrea en el dorso.
Anteras blanquecinas, sin polen. Ramas estilares amarillas, en ocasiones tenidas de púrpura. Aquenios maduros desconocidos,
los inmaduros de color pajizo. Vilano 5–6 mm de longitud, discoloro, blanco, verdoso en la base.
Hábitat y distribución.—Solo se conoce un único testimonio de herbario —BR 970947— que contiene una etiqueta con una descripción geográfica poco precisa.
No se ha podido volver a recolectar. Habita en pastizales húmedos del pirineo leridano, alrededor de 1200 m (fig. 6).
|
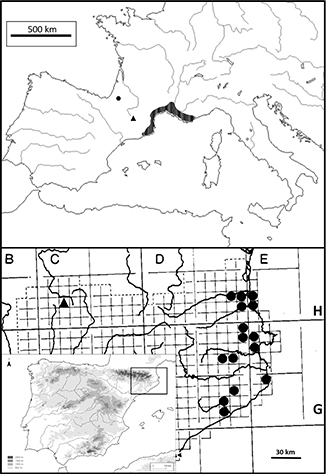 Fig. 6. Distribución de Taraxacum litophyllum De Langhe & Soest [triángulo negro, localización aproximada; BR 970947] y T. raii (Gouan) S.F.Gray en Europa y la península ibérica [círculos negros, localidades aisladas; tramado, área continua]. [Ambos
mapas están elaborados con datos propios, de herbario y bibliográficos (Soest 1961, 1965; Krischner & Štĕpánek 1998; Molina 2015; Noble & al. 2016; Tison 2017).] Fig. 6. Distribución de Taraxacum litophyllum De Langhe & Soest [triángulo negro, localización aproximada; BR 970947] y T. raii (Gouan) S.F.Gray en Europa y la península ibérica [círculos negros, localidades aisladas; tramado, área continua]. [Ambos
mapas están elaborados con datos propios, de herbario y bibliográficos (Soest 1961, 1965; Krischner & Štĕpánek 1998; Molina 2015; Noble & al. 2016; Tison 2017).]
|
|
Fenología.—La única planta recolectada estaba florecida en julio.
Conservación.—Se dispone de muy pocos datos sobre este taxón, pero lo suponemos raro y, de momento, endémico del NE de la península ibérica.
Hace falta prospectar mucho más el Parque Nacional de Aigües Tortes i Estany de Sant Maurici y sus alrededores para conocer
el estado de su conservación, así como para completar su descripción botánica. Actualmente no se le puede otorgar ningún estatus
de conservación IUCN concreto, y se ha de considerar como un taxón con datos insuficientes —DD.
Observaciones.—Taraxacum litophyllum podría confundirse con T. ciliare, como sugiere la combinación T. palustre subsp. litophyllum, pues T. palustre, aunque no está en la península, es cercano a T. ciliare; sin embargo, T. litophyllum florece en verano, presenta pecíolos alados, las flores no contienen polen, la corola es azafranada, y el vilano discoloro.
Material estudiado.—ESPAÑA. Lleida: Parque Nacional de Aigües Tortes, 1200 m, 8-VII-1970, J. De L. [Lange] (BR 970947).
4. T. raii (Gouan) S.F.Gray, Nat. Arr. Brit. Pl. 2: 426 (1821). Leontodon raii Gouan, Ill. Observ. Bot.: 55 (1773), basón. Tipo: Francia, [Hérault], Montpellier, Villeneuve, 23-II-1913, J. Braun y E. Furrer s.n. (neo-, designado por Kirschner & Štĕpánek (1998: 21): Z 3949!). Fig. 7.
|
 Fig. 7. Taraxacum raii (Gouan) S.F.Gray: a, hábito; b, capítulo en flor con las brácteas externas adpresas; c, variabilidad de brácteas externas —las dos de la izquierda— e internas —las dos de la derecha—; d, polen junto a los tejidos de una antera; e, polen del tamaño regular; f-h, variabilidad del cuerpo y del cono de los aquenios; i-l, variabilidad de las hojas [a, j, HGI 23429; b-c, HGI 23433; d, HGI 18897; e, HGI 18996; f, HGI 23417; g, HGI 23426; h, HGI
8627; i, HGI 23428; k, HGI 23419; l, HGI, 23423]. Fig. 7. Taraxacum raii (Gouan) S.F.Gray: a, hábito; b, capítulo en flor con las brácteas externas adpresas; c, variabilidad de brácteas externas —las dos de la izquierda— e internas —las dos de la derecha—; d, polen junto a los tejidos de una antera; e, polen del tamaño regular; f-h, variabilidad del cuerpo y del cono de los aquenios; i-l, variabilidad de las hojas [a, j, HGI 23429; b-c, HGI 23433; d, HGI 18897; e, HGI 18996; f, HGI 23417; g, HGI 23426; h, HGI
8627; i, HGI 23428; k, HGI 23419; l, HGI, 23423].
|
|
T. ruborum Soest, Acta Bot. Neerl. 10: 286 (1961)
T. brachysquameum Soest, Acta Bot. Neerl. 14: 23 (1965)
Hierba hasta de 40 cm. Tallos erectos, tan largos o más que las hojas, sin hojas bracteiformes, pubescentes, con indumento
aracnoideo, lampiños al madurar, quedando en ocasiones algo de pilosidad bajo el capítulo, verdes, pardos o purpúreos. Hojas
de 9–23 × 0,6–3,5 cm, pecioladas, de contorno muy variable, espatulado-lanceoladas, ligeramente dentadas o de pinnatífidas
a pinnatisectas, suaves, verdes, sin manchas purpúreas; nervio medio sin estrías longitudinales, verde o purpureo; lóbulos
laterales 0–6 por hemilimbo, de 2,0–20,1 × 6,1–25,2 mm —relación entre el largo y el ancho 0,2–0,9—, de triangulares a deltoideos,
indivisos, con el margen distal entero, recto o sigmoideo y el proximal entero, recto o ligeramente sigmoideo, con los dientes
de 1–2 mm, triangulares, con o sin dientes entre los lóbulos, en ocasiones los proximales se continúan en el pecíolo con algún
diente triangular; lóbulo terminal 8,8–48,7 × 70,0–36,0(47,0) mm —relación entre el largo y el ancho 0,9–2,2—, generalmente
agudo, de forma variable, la estirpe típica presenta frecuentemente forma de obús, con o sin dientes distantes y los márgenes
de la base simétricos o asimétricos; pecíolo de verde a purpureo, estriado longitudinalmente, áptero, sin dientes. Capítulo
hasta de 25(30) mm de diámetro en la antesis. Involucro 12–20 × 6–9 mm; brácteas externas 3,4–9,0 × 2,3–5,2 mm, normalmente
ovado-lanceoladas, más raramente ovadas —relación entre el largo y el ancho de 1,4–2,4—, adpresas o erectas, a veces de ligeramente
recurvadas a patentes durante la madurez, con margen escarioso de 0,1–0,3(0,6) mm de anchura, de verde claro a verde obscuro,
muy a menudo teñidas de púrpura hacia los márgenes y el ápice, cilioladas, sin cuerno o callo subapical. Flores hermafroditas.
Corolas hasta de 20 mm de longitud, amarilla, las más externas del capítulo con una banda ancha rojiza en el dorso. Anteras
amarillas, con granos de polen de tamaño regular o algunos pocos de distintos tamaños. Ramas estilares frescas de amarillo
a verde amarillento más o menos pálido, de amarillo negruzco a negro cuando secas. Aquenios de un verde amarillento, verde
oliva o parduscos; cuerpo (2,3)3,0–3,9 mm de longitud, con escuámulas erosas, bi o tricuspidadas y espículos cortos hacia
el ápice, el resto liso, con las costillas ± anchas; cono (0,4)0,5–0,8(0,9) mm de longitud, de cilíndrico a cónico; pico (2,8)5,0–9,5(10,5)
mm, blanquecino. Vilano 3,5–7,0 mm de longitud, concoloro, blanco. 2n = 16.
Hábitat.—Habita en prados y juncales higrófilos, principalmente en prados de siega temporalmente inundados —Trifolio-Hordeetalia—, prados litorales subsalinos de Carex divisa y tréboles marinos —Juncion maritimi, Trifolion maritimi— y herbazales de Oenanthe fistulosa —Ranunculo-Oenanthion—. También se puede encontrar esporádicamente en fresnedas inmaduras —Carici-Fraxinion oxycarpae, Fraxino-Ulmenion Rivas-Martínez 1975— que corresponden a antiguos prados de siega abandonados (Donker & Stevelink 1961; Mercadal & al. 2008; Mercadal & Vilar 2013). Los suelos son preferentemente arcillosos y normalmente paraturbosos, de reacción indiferente, incluso subsalinos, y siempre
temporalmente inundados (tabla 3). Altitudinalmente se distribuye de 0–200 m.
Tabla 3. Características edáficas de los prados donde habita Taraxacum raii (Gouan) S.F.Gray. [ISSS, International Society of Soil Science.]
| Valores |
pH 1:2,5 H2O
|
CE 1:5 a 25ºC (µS/cm) |
Carbonatos (%) |
Materia orgánica (%) |
Humedad de campo (%) |
Clase textural [ISSS] (número de muestras) |
| Mínimo |
6,5 |
77,4 |
0,0 |
1,7 |
20,1 |
arcillosa fina (5) |
| Máximo |
8,3 |
1022,7 |
6,0 |
4,5 |
38,5 |
arcillosa gruesa (2) |
| Media |
7,4 |
307,9 |
1,0 |
3,0 |
29,1 |
arcillosa limosa (1) |
| Desviación estándar |
0,6 |
276,0 |
1,8 |
0,9 |
4,8 |
arcillosa arenosa (1) |
| Número de muestras |
21 |
21 |
11 |
11 |
21 |
franco arcillosa (1) |
Fenología.—De marzo a mayo.
Distribución.—Área litoral y prelitoral del suroeste de Europa: sur de Francia, en Languedoc —locus classicus—, Provenza y, puntualmente, en el departamento de los Pirineos Atlánticos (Soest 1961, 1965; Krischner & Štĕpánek 1998; Molina 2015; Noble & al. 2016; Tison 2017) y NE de España. En la península ibérica se ha herborizado hasta ahora en el NE de Cataluña, en la comarca litoral barcelonesa
del Maresme, y en las gerundenses del Alt Empordà, del Baix Empordà, del Gironès, del Pla de l’Estany y de la Selva. Más al
norte, también se ha colectado en el Rosselló, en el S de Francia. Además, cabe esperar que pueda extenderse más hacia el
sur, seguramente hasta el Delta del Llobregat, donde hay hábitats apropiados para su desarrollo (fig. 6).
Conservación.—Kirschner & Štĕpánek (1998) consideraron este taxón como endémico —EN— del S de Francia, pero T. raii penetra como mínimo hasta el noreste de la península ibérica. Por otra parte, Molina (2015) y Noble & al. (2016) no le asignan ningún estatus de conservación IUCN por poseer pocos datos, y lo consideran con datos insuficientes —DD— en
la región Languedoc-Roussillon y en la région Provence-Alpes-Côte d’Azur respectivamente. A nuestro entender, su estatus más
idóneo, a partir del área de distribución conocida hasta ahora, sería el de casi amenazado —NT—, por poseer varios núcleos
poblacionales pero que se desarrollan en zonas húmedas más o menos amenazadas por el saneamiento y la urbanización.
Observaciones.—Malato-Beliz (1973) citó T. ruborum en el distrito de Portalegre —Portugal—, pero no hemos encontrado ningún testimonio de herbario que confirme esta observación.
La planta fue vista a 600 m de altitud, 400 m por encima de cualquier población de T. raii observada hasta ahora, y a una distancia considerable del mar, lo que hace sospechar que ha habido un error en la identificación
que tendría que ser confirmado con el estudio del correspondiente testimonio. Taraxacum raii se distingue bien del resto de congéneres ibéricos de T. sect. Palustria por poseer los granos de polen de tamaño regular o casi regular —< 10% de los granos son desiguales— al observarlos a 600×.
Por lo tanto, junto con T. tenuifolium (Hoppe & Hornsch.) W.D.J.Koch, Flora 23: 373–374 (1840), de Italia y la antigua Yugoslavia, es uno de los dos taxones diploides
de la sección en Europa que presentan reproducción sexual (Kirschner & al. 2017). En cambio, el hecho de presentar los estigmas y estilos ennegrecidos una vez la planta se encuentra seca no es una característica
discriminante tal y como sugirió Soest (1965); por ejemplo, T. aginnense, T. ciliare y T. gr. mediterraneum también pueden presentar a veces los estigmas y estilos muy ennegrecidos.
Material estudiado.—ESPAÑA. Barcelona: Tordera, al pla d’Anyells, prat de can Gener, 31TDG7618, 32 m, 19-III-2017, G. Mercadal (HGI 23508); Tordera, a l’estany de can Torrent, 31TDG7516, 24 m, 19-III-2017, G. Mercadal (HGI 23529). Girona: Banyoles, marges de l’Estany, vora la font de la Filadora, 31TDG7962, 174 m, 30-IV-2015, G. Mercadal (HGI 22710); Caldes de Malavella, als prats de Sant Sebastià, 31TDG8130, 79 m, 11-IV-2015, G. Mercadal (HGI 22695); Caldes de Malavella, als prats de Sant Sebastià, 31TDG8130, 79 m, 17-IV-2015, G. Mercadal (HGI 22706); Cantallops, al Bell-lloc, 31TDG9494, 142 m, 8-V-2004, G. Mercadal (HGI 18998); Capmany, a Querafumat, 31TEG9194, 148 m, 11-IV-2003, G. Mercadal (HGI 18996); Castelló d’Empúries, a les closes de la Paborderia, 31TEG0679, 4 m, 12-IV-2016, G. Mercadal (HGI 23429); La Jonquera, al prat Llong dels estanys del Canadal, 31TDG9194, 178 m, 22-III-2003, G. Mercadal (HGI 18997), La Jonquera, al prat Llarg de Canadal, 31TDG9194, 177 m, 31-III-2016, G. Mercadal (HGI 23419); Llagostera, ca n’Oms, 31TDG8833, 108 m, 2-IV-2016; G. Mercadal (HGI 23417); Pals, a les closes Fondes, 31TEG1349, 1 m, 12-IV-2016, G. Mercadal (HGI 23426); Pals, a les closes Fondes, 31TEG1349, 1 m, 13-IV-2016, G. Mercadal (HGI 23428); Peralada, a les closes de l’Ullal, 31TEG0781, 2 m, 15-IV-2015, G. Mercadal (HGI 22707); Rabós, can Costoja, 31TEG0388, 49 m, 26-III-2016, G. Mercadal (HGI 23425); Sant Climent Sescebes, a les Closes, 31TDG9592, 125 m, 4-IV-1995, J. Font (HGI 8627); Sant Climent Sescebes, riera de Forns, 31TDG9689, 83 m, 24-IV-2015; G. Mercadal (HGI 22719); Sils, a l’antic estany, 31TDG7627, 65 m, 20-III-2008, G. Mercadal (HGI 22105).
FRANCIA. Pyrénées-Orientales: Argelers, les Conques, 31TEH0211, 3 m, 31-III-2016, G. Mercadal (HGI 23434); Cànoes, a l’antic estany, 31TDH8722, 55 m, 17-IV-2015, G. Mercadal (HGI 22708); Sant Cebrià, prats de l’Aigual, 31TEH0119, 2 m, 31-III-2015, G. Mercadal (HGI 23433); Sant Cebrià: prats de l’Aigual, 31TEH0119, 2 m, 31-III-2015, G. Mercadal (HGI 23432); Sant Nazari, a l’antic estany, 31TEH0024, 1 m, 31-III-2015, G. Mercadal (HGI 23430); Sant Nazari, a l’antic estany, 31TDH9923, 1m, 31-III-2016, G. Mercadal (HGI 23431); Tuïr, a la Prada, 31TDH8120, 85 m, 7-IV-2016; G. Mercadal (HGI 23423); Tuïr, a la Prada, 31TDH8121, 85 m, 7-IV-2016, G. Mercadal (HGI 23424); Tuïr, a la Prada, 31TDH8120, 85 m, 17-IV-2015, G. Mercadal (HGI 22705).
Taraxacum sect. Erythrocarpa Hand.-Mazz.
5. T. gr. mediterraneum [incl.: T. mediterraneum Soest, Vegetatio 5–6: 530 (1954); T. balearicum Soest, Acta Bot. Neerl. 10: 282 (1961); T. mediterraneiforme Soest, Acta Bot. Neerl. 10: 285 (1961); T. dargilanicum Sonck, Ann. Bot. Fennici 22: 251 (1985)]. Fig. 8.
|
 Fig. 8. Taraxacum gr. mediterraneum: a, hábito; b, hojas; c, capítulo con las brácteas externas patentes; d, variabilidad de las brácteas externas; e, f, variabilidad del cuerpo y cono de los aquenios; g, polen de tamaño irregular junto a los tejidos de una antera [a-d, HGI 23444; e, f, HGI 23439; g, HGI 23440]. Fig. 8. Taraxacum gr. mediterraneum: a, hábito; b, hojas; c, capítulo con las brácteas externas patentes; d, variabilidad de las brácteas externas; e, f, variabilidad del cuerpo y cono de los aquenios; g, polen de tamaño irregular junto a los tejidos de una antera [a-d, HGI 23444; e, f, HGI 23439; g, HGI 23440].
|
|
Hierba hasta de 50 cm. Tallos erectos, más cortos o, habitualmente, más largos que las hojas, sin hojas bracteiformes, pubescentes,
con indumento aracnoideo, lampiños al madurar, quedando en ocasiones algo de pelosidad bajo el capítulo, verdes con tonalidades
purpúreas. Hojas 2,7–26,2 × 0,7–5,8 cm, pecioladas, espatuladas, dentadas y más ordinariamente de pinatífidas a pinnatisectas,
suaves, verdes, sin manchas purpúreas; nervio medio sin estrías longitudinales, de verde a purpureo; lóbulos laterales 2–7
por hemilimbo, de 3,5–15,1 × 3,1–22,2 mm —relación entre el largo y el ancho 0,4–1,5—, deltoideos, indivisos, con el margen
distal entero, recto o ligeramente sigmoideo, y el proximal entero, de recto a sigmoideo, a veces con dientes triangulares
de 1,4–3,6 mm entre los lóbulos, los proximales se continúan en el pecíolo, a veces con dientes triangulares o filiformes;
lóbulo terminal 9,1–32,0 × 7,8–32,0 mm —relación entre el largo y el ancho (0,9)1,1–1,7—, de triangular a sagitado, con el
ápice redondeado, a veces mucronulado, entero, con los márgenes de la base frecuentemente asimétricos; pecíolo verde, sin
estrías longitudinales, áptero, sin dientes. Capítulo de 18–43 mm de diámetro en la antesis. Involucro 12,0–22,3 × 3,9–10,0
mm; brácteas externas 2,9–10,2 × 2,3–4,4 mm, de lanceoladas a ovado-lanceoladas —relación entre el largo y el ancho (1,9)2,2–2,9(3,4)—,
erectas, patentes o ligeramente recurvadas, con margen escarioso 0,1–0,4 mm de anchura, en ocasiones inconspicuo, verdes,
ligeramente purpúreas hacia los márgenes y el ápice, a veces cilioladas, en ocasiones con un callo subapical. Flores hermafroditas.
Corolas hasta de 21,0 mm de longitud, amarilla, las más externas del capítulo con una banda ancha de color violeta obscuro
en el dorso. Anteras amarillas, con granos de polen de distintos tamaños. Ramas estilares de color amarillo a verde amarillento,
de marrón obscuro a negro cuando secas. Aquenios de color verde oliva a parduscos; cuerpo (2,2)2,8–3,8(4,0) mm de longitud,
con papilas y espículos cortos y gruesos hacia el ápice, el resto liso, con las costillas estrechas; cono 0,4–0,8 mm de longitud,
de subcilíndrico a cónico; pico (4,4)5,5–7,6(8,8) mm de longitud, de blanco a verdoso. Vilano 4,5–7,0(8,0) mm de longitud,
concoloro, blanco. 2n = 24, 32.
Hábitat.—Habita en márgenes de caminos, zonas umbrías de jardines, fenalares —Brachypodion phoenicoidis Br.-Bl. ex Molinier 1934—, prados de siega higrófilos —Trifolio-Hordeetalia— y hasta en herbazales húmedos —Ranunculo-Oenanthion—. Resiste el pastoreo temporal. En la península hemos recolectado los especímenes en el piso basal, de 0–600 m; pero Girerd & Roux (2011) indican una población hasta los 1500 m, en el Mont Ventoux —Provenza—. Los suelos son arcillo-arenosos, de reacción neutra
o ligeramente básica, a veces subsalinos, y con la humedad de campo muy variable, de mesófilos —lo más común— a higrófilos
(tabla 4).
Tabla 4. Características edáficas de los prados donde habita Taraxacum gr. mediterraneum. [ISSS, International Society of Soil Science.]
| Valores |
pH 1:2,5 H2O
|
CE 1:5 a 25ºC (µS/cm) |
Carbonatos (%) |
Materia orgánica (%) |
Humedad de campo (%) |
Clase textural [ISSS] (número de muestras) |
| Mínimo |
6,2 |
56,5 |
0,0 |
2,5 |
15,6 |
arcillosa gruesa (1) |
| Máximo |
7,8 |
1220,4 |
1,3 |
2,9 |
29,0 |
franco arenosa (1) |
| Media |
6,8 |
366,6 |
0,9 |
2,7 |
23,8 |
— |
| Desviación estándar |
0,7 |
570,0 |
0,7 |
0,2 |
5,9 |
— |
| Número de muestras |
4 |
4 |
3 |
3 |
4 |
— |
Fenología.—De marzo a mayo.
Distribución.—Región mediterránea occidental, S de Francia y NE de España: desde el departamento de los Alpes marítimos —Provenza— hasta
el noreste de Barcelona —Cataluña— y Mallorca —Islas Baleares—(Soest 1954, 1961; Sonck 1985; Pla & al. 1992; Girerd & Roux 2011; Tison & al. 2014; Molina 2015; Galán de Mera & Sáez 2016; Noble & al. 2016). En la península ibérica se ha herborizado hasta ahora en el noreste de Cataluña, en la comarca litoral barcelonesa del
Maresme, y en las gerundenses del Alt Empordà y de la Selva. Más al norte, también se ha colectado en el Rosselló —Francia—
(fig. 9).
|
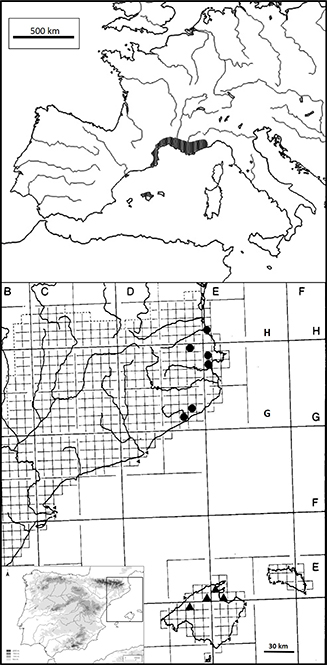 Fig. 9. Distribución de Taraxacum gr. mediterraneum en Europa y la península ibérica [círculos negros, localidades aisladas; tramado, área continua] y de T. mediterraneum Soest s.str. en las Islas Baleares [triángulos negros, localidades aisladas]. [Mapa del NE de la Península, S de Francia
e Islas Baleares elaborado con datos propios; el de Europa con referencias bibliográficas (Soest 1954, 1961; Sonck 1985; Girerd & Roux 2011; Molina 2015; Noble & al. 2016).] Fig. 9. Distribución de Taraxacum gr. mediterraneum en Europa y la península ibérica [círculos negros, localidades aisladas; tramado, área continua] y de T. mediterraneum Soest s.str. en las Islas Baleares [triángulos negros, localidades aisladas]. [Mapa del NE de la Península, S de Francia
e Islas Baleares elaborado con datos propios; el de Europa con referencias bibliográficas (Soest 1954, 1961; Sonck 1985; Girerd & Roux 2011; Molina 2015; Noble & al. 2016).]
|
|
Conservación.—En la península ibérica aún se dispone de poca información corológica; por esta razón, provisionalmente, se considera como
un taxón con datos insuficientes —DD—, como opinan Molina (2015) y Noble & al. (2016) para las regiones Languedoc-Roussillon y Provence-Alpes-Côte d’Azur respectivamente. Aun así, en su área global, T. gr. mediterraneum presenta una distribución compacta desde la Provenza a Cataluña e Islas Baleares, así como una ecología amplia, por lo tanto
se estima como de preocupación menor —LC.
Observaciones.—Bajo el nombre T. gr. mediterraneum incluimos un conjunto microespecies —T. mediterraneum, T. mediterraneiforme, endémico de Provenza, y T. dargilanicum, endémico de Languedoc—difíciles de delimitar por su plasticidad morfológica (Tison & al. 2014), pero que presentan unas características comunes bien distinguibles del resto de congéneres próximos de T. sect. Erythrocarpa, T. sect. Celtica A.J.Richards y T. sect. Palustria. Sin embargo, respecto a nuestras poblaciones, T. mediterraneiforme es de menor talla —c. 10 cm—, presenta el pecíolo purpúreo y 4–5 lóbulos laterales agudos de 15–20 mm de longitud (Soest 1961); T. dargilanicum reúne plantas de 10–15 cm, con el pecíolo purpúreo y 3–4 lóbulos laterales agudos de 10–20 mm de longitud (Sonck 1985). Los especímenes estudiados en el NE de la Península y SE de Francia presentan algunos caracteres diferenciales frente al
T. mediterraneum s.str. recolectado en Baleares (Galán de Mera & Sáez 2016): hojas de mayor tamaño —2,7–12 × 0,7–2,1 cm en T. mediterraneum—, capítulos más grandes —c. 20 mm en T. mediterraneum—, brácteas externas de mayores dimensiones —2,9–5,9 × 2,3–3,2 mm en T. mediterraneum— y nunca con un callo subapical. Por ello no incluimos este material en ninguna de las citadas microespecies. Tanto las plantas
catalanas como baleáricas son poliploides y su polen irregular sugiere la agamospermia como comportamiento reproductivo (Uhlemann 2001). Tras estudiar el material de T. mediterraneum y T. balearicum conservado en BR, recolectado en Mallorca por J. Duvigneaud, y la mayoria determinado por J.L. van Soest (Duvigneaud 1979), hemos llegado a la conclusión de que son sinónimos. Todos los ejemplares presentan las hojas con los lóbulos laterales
deltoideos y asimétricos, el involucro con las brácteas externas de adpresas a recurvadas, y los aquenios de amarillentos
a parduscos con el cono subcilíndrico. Por otra parte, en el holotipo de T. balearicum (L 0002367!) las hojas aparecen poco conservadas y faltan los aquenios, por lo que solo podemos comparar con los parátipos
de BR. Los lóbulos asimétricos de las hojas recuerdan a T. vinosum Soest, Collect. Bot. (Barcelona) 4: 29 (1954) —T. sect. Erythrosperma—, pero los pecíolos verdes, las brácteas externas del involucro menores, la presencia de polen en las flores, y el color y
la estructura de los aquenios diferencian claramente a T. gr. mediterraneum de T. vinosum. Por último, no hemos podido confirmar la cita aragonesa de T. balearicum a la que se refiere Villar & al. (2001), aunque parece dudosa.
Material estudiado.—ESPAÑA. Barcelona: Tordera, a l’estany de can Torrent, 31TDG7516, 24 m, 19-III-2017, G. Mercadal (HGI 23524); Tordera, a l’estany de can Torrent, 31TDG7516, 24 m, 19-III-2017, G. Mercadal (HGI 23524). Girona: Agullana, prats de Santa Eugènia, 31TDG8595, 312 m, 7-IV-2016, G. Mercadal (HGI 23439); Peralada, a les closes dels Gaus, 31TEG0183, 18 m, 12-IV-2016, G. Mercadal (HGI 23443); Sils, als Tres Ponts, 31TDG7928, 67 m, 15-IV-2017, G. Mercadal (HGI23523); Vajol, prats del Perdigó, 31TDG8294, 542 m, 13-IV-2016, G. Mercadal (HGI 23444). Illes Balears-Mallorca: Chemin à l’entrée du jardin d’Alfabia (environs du col de Sóller), 2-IV-1913, L. Kofler (L 0002367, holotipo de T. balearicum); L’Alfabia, en las jardineras del palacio, 31SDD79, 219 m, 30-IV-2007, A. Galán 1868 (MA 804779); ibid., A. Galán 1869 (USP 3226); Artà, ermita de Betlem, bord de chemin vers la fontaine, 31SEE10, 29 m, 18-IV-1974, J. Duvigneaud 74E269 (BR 618337); Lluch, cour du monastère, 31SDE90, 500 m, 13-IV-1971, J. Duvigneaud 71E101 (BR 618350, como T. cupreum); ibid., groupements herbeux sous les arbres, 11-IV-1965, J. Duvigneaud 65E186 (BR 16138445, como T. retzii); Pollensa, 31SEE01, 59 m, 20-IV-1973, J. Duvigneaud 73E193 (BR 618356, como T. balearicum).
FRANCIA. Pyrénées-Orientales: Sant Cebrià, prats de l’Aigual, 31TEH0119, 2 m, 31-III-2016, G. Mercadal (HGI 23440).
AGRADECIMIENTOSTOP
Deseamos expresar nuestro agradecimiento al Jardín Botánico de Meise —Agentschap Plantentuin Meise— por facilitarnos la imagen del pliego de T. litophyllum —BR 970947— y permitir su reproducción; al Sr. J.-M. Tison, de la Société Botanique de France, por la confirmación y comentarios sobre algunos testimonios de herbario complejos.
REFERENCIASTOP
| ○ |
Aquaro G., Caparelli K.F. & Peruzzi L. 2008. The genus Taraxacum (Asteraceae) in Italy. I. A systematic study of Taraxacum sect. Palustria. Phytologia Balcanica 14(1): 61–67.
|
| ○ |
Dahlstedt H. 1921. De Svenska Arterna av Släktet Taraxacum. Acta Florae Sueciae 1: 1–160.
|
| ○ |
De Langhe J.E. & Soest J.L. van 1972. Deux Taraxacum noveaux découverts dans les Pyrénées et en Auvergne: Taraxacum litophyllum De Langhe et van Soest et Taraxacum arvernum De Langhe et van Soest. Bulletin de la Société Royale de Botanique de Belgique 105(1): 115–118.
|
| ○ |
Donker M. & Stevelink A. 1961. Einige Wiesenvegetationen (Gaudinio-Arrhenatheretum; Molinietum mediterraneum; Caricetum divisae) im Vistre-Tal bei Le Cailar. Meded Landbouwh Wageningen 61(15): 1–32.
|
| ○ |
Duvigneaud J. 1979. Catalogue provisoire de la flore des Baléares. Bulletin de la société pour l’échange des plantes vasculaires de l’Europe occidentale et du bassin méditerranéen 17 (Suppl.): 1–43.
|
| ○ |
Galán de Mera A. 2017. Taraxacum F.H. Wigg. In Talavera S., Buira A., Quintanar A., García M.A., Talavera M., Fernández Piedra T. & Aedo C. (eds.), Flora iberica 16 (2): 963–1062. CSIC, Madrid.
|
| ○ |
Galán de Mera A. & Sáez L. 2016. Taraxacum majoricense (Asteraceae), a new species from the Balearic Islands, Spain. Annales Botanici Fennici 53: 82–90. https://doi.org/10.5735/085.053.0216 |
| ○ |
Girerd B. & Roux J.-P. 2011. Flore du Vaucluse. Troisième inventaire, descriptif, écologique et chorologique. Biotope, Mèze.
|
| ○ |
Hawksworth D.L. 2002. The Changing Wildlife of Great Britain and Ireland. CRC Press, Nueva York.
|
| ○ |
Hofstra J. 1985. Morphologie, phytosociologie et écologie de quelques espèces de Taraxacum sect. Palustria Dahlstedt en Agenais (France). Bulletin de la Société Botanique de France 132: 123–131. http://dx.doi.org/10.1080/01811797.1985.10824659 |
| ○ |
Hofstra J. 1990. L’Orchio-Brometum du Lot-et-Garane. Bulletin de la Société d’histoire naturelle de Toulouse 126: 79–84.
|
| ○ |
Kirschner J. & Štĕpánek J. 1998. A monograph of Taraxacum sect. Palustria. Academy of Sciences of the Czech Republic, Praga.
|
| ○ |
Kirschner J., Štĕpánek J. & Greuter W. 2017. Taraxacum. In Greuter W. & Raab-Straube E. von (eds.), Compositae. Euro+Med Plantbase – the information resource for Euro-Mediterranean plant diversity. Página web: http://ww2.bgbm.org/EuroPlusMed/query.asp [consultada noviembre 2016].
|
| ○ |
Kirschner J., Štěpánek J., Mes T.H.M., Den Nijs J.C.M., Oosterveld P., Štorchová H. & Kuperus P. 2003. Principal features
of the cpDNA evolution in Taraxacum (Asteraceae, Lactuceae): a conflict with taxonomy. Plant Systematics and Evolution 239: 231–255. https://doi.org/10.1007/s00606-003-0002-5 |
| ○ |
Kirschner J., Záveská Drábková L., Štěpánek J. & Uhlemann I. 2015. Towards a better understanding of the Taraxacum evolution (Compositae-Cichorieae) on the basis of nrDNA of sexually reproducing species. Plant Systematics and Evolution 301: 1135–1156. https://doi.org/10.1007/s00606-014-1139-0 |
| ○ |
Lambinon J. & Verloove F. 2012. Nouvelle Flore de la Belgique, du Gran-Duché de Luxembourg, du Nord de la France et des Régions voisines (Ptéridophytes et
Spermatophytes), ed. 6. Edition du Jardin botanique de Belgique, Meise.
|
| ○ |
Malato-Beliz J. 1973. Novas especies para a flora de Portugal, II. Lagascalia 3 (1): 61–69.
|
| ○ |
McNeill J., Barrie F.R., Buck W.R., Demoulin V., Greuter W., Hawksworth D.L., Herendeen P.S., Knapp S., Marhold K., Prado
J., Prud’homme van Reine W.F., Smith G.F., Wiersema J.H. & Turland N.J. 2012. International Code of Nomenclature for algae, fungi, and plants (Melbourne Code). Koeltz Scientific Books, Koenigstein.
|
| ○ |
Mercadal G. & Vilar L. 2013. Caracterització de les freixenedes al·luvials inundables del nord-est de Catalunya (Carici remotae-Fraxinetum oxycarpae Pedrotti 1970 corr. Pedrotti 1992). Orsis 27: 53–94.
|
| ○ |
Mercadal G., Gesti J., Corominas M. & Vilar L. 2003. Los prados de guadaña de la región mediterránea de Catalunya. Pastos 33 (2): 219–232.
|
| ○ |
Mercadal G., Vilar L. & Gesti J. 2008. L’herbassar de felandri fistulós i baldèl·lia (Baldellio ranunculoidis-Oenanthetum fistulosae), una nova associació herbàcia higròfila dels Països Catalans. Orsis 23: 47–73.
|
| ○ |
Molina J. (coord.). 2015. Catalogue de la flore vasculaire de la région Languedoc-Roussillon, v. 2. Página web: http://bdd.flore.silene.eu/catalogue_reg/lr/index.php [consultada noviembre 2016].
|
| ○ |
Noble V., Van Es J., Michaud H. & Garraud L. (coords.). 2016. Catalogue de la flore vasculaire de la région Provence-Alpes-Côte
d’Azur, v. 2.4. Página web: http://bdd.flore.silene.eu/catalogue_reg/paca/index.php [consultada febrero 2017].
|
| ○ |
Pla V., Sastre B. & Llorens L. 1992. Aproximació al catàleg de la flora vascular de les Illes Balears. Universitat de les
Illes Balears et Jardí Botànic de Sóller, Palma de Mallorca.
|
| ○ |
Porta J., López-Acevedo M. & Rodríguez R. 1986. Técnicas y experimentos en edafología. Col·legi Oficial d’Enginyers Agrònoms de Cataluña, Lleida.
|
| ○ |
Porta J., López-Acevedo M. & Roquero C. 2003. Edafología para la agricultura y el medio ambiente, ed. 3. Mundi-Prensa, Madrid.
|
| ○ |
Richards A.J. & Haworth C.C. 1984. Further new species of Taraxacum from the British Isles. Watsonia 15: 85–94.
|
| ○ |
Richards A.J. & Sell P.D. 1984. Taraxacum Weber. In Tutin T.G. & al. (eds.), Flora Europaea 4: 332–343. Cambridge University Press, Cambridge.
|
| ○ |
Scopoli A. 1772. Flora carniolica ed. 2, vol. 2. Joannis Thomae Trattner, Viena.
|
| ○ |
Sell P. & Murrell G. 2006. Flora of Great Britain and Ireland, 4. Campanulaceae-Asteraceae. Cambridge University Press, Nueva York.
|
| ○ |
Soest J.L. van 1954. Sur quelques Taraxaca d’Espagne. Collectanea Botanica (Barcelona) 4: 1–32.
|
| ○ |
Soest J.L. van 1961. Quelques nouvelles espèces de Taraxacum, natives d’Europe. Acta Botanica Neerlandica 3 (10): 280–306. https://doi.org/10.1111/j.1438-8677.1961.tb00055.x |
| ○ |
Soest J.L. van 1965. Taraxacum sect. Palustria Dahlstedt. Acta Botanica Neerlandica 14 (1): 1–53. https://doi.org/10.1111/j.1438–8677.1965.tb00178.x |
| ○ |
Soldano A. 2010. Notulae alla checklist della flora vascolare italiana 9 (1623–1681). Informatore Botanico Italiano 42 (1): 369–389.
|
| ○ |
Sonck C.E. 1985. Espèces nouvelles de Taraxacum de France II. Annales Botanici Fennici 22: 245–253.
|
| ○ |
Thiers B. 2017. Index Herbariorum: a global directory of public herbaria and associated staff. Página web: http://sweetgum.nybg.org/ih/ [consultada noviembre 2016].
|
| ○ |
Tison J.-M. 2017. Photoflora. Base de données de photos de plantes essentiellement de la flore européenne Página web: http://photoflora.free.fr/index.php [consultada marzo 2017].
|
| ○ |
Tison J.-M. & Foucault B. 2014. Flora Gallica. Biotope, Mèze.
|
| ○ |
Tison J.-M., Jauzein P. & Michaud H. 2014. Flore de la France méditerranéenne continentale. Naturalia, Turriers.
|
| ○ |
Uhlemann I. 2001. Distribution of reproductive systems and taxonomical concepts in the genus Taraxacum F.H. Wigg. (Asteraceae, Lactuceae) in Germany. Feddes Repertorium 112: 15–35. https://doi.org/10.1002/fedr.20011120105 |
| ○ |
Uribe-Echebarría P.M. 1999. Taraxacum Weber. In Aizpuru I. & al. (eds.), Claves ilustradas de la flora del País Vasco: 573–574. Gobierno Vasco, Vitoria-Gasteiz.
|
| ○ |
Vašut R.J. 2003. Taraxacum sect. Erythrosperma in Moravia (Czech Republic): Taxonomic notes and the distribution of previously described species. Preslia 75: 311–338.
|
| ○ |
Villar L., Sesé J.A. & Ferrández J.V. 2001. Atlas de la Flora del Pirineo Aragonés, vol. 2 (Pyrolaceae-Orchidaceae). Consejo de Protección de la Naturaleza de Aragón & Instituto de Estudios Altoaragoneses, Huesca.
|
| ○ |
Watanabe K. 2017. Index of Chromosome Numbers in Asteraceae. Página web: http://www.lib.kobe-u.ac.jp/infolib/meta_pub/G0000003asteraceae_e [consultada noviembre 2016].
|
| ○ |
Willkomm M. & Lange J. 1865. Taraxacum Juss. In Willkomm M. & Lange J., Prodromus Florae Hispanicae 2 (1): 230–231. E. Schweizerbart (E. Koch), Estucardia.
|
| ○ |
Wittzell H. 1999. Chloroplast DNA variation and reticulate evolution in sexual and apomíctic sections of dandelions. Molecular Ecology 8: 2023–2035. https://doi.org/10.1046/j.1365-294x.1999.00807.x |
| ○ |
Záveský L., Jarolímova V. & Štěpánek J. 2007. Apomixis in Taraxacum paludosum (section Palustria, Asteraceae): Recombinations of apomixis elements in inter-sectional crosses. Plant Systematics and Evolution 265: 147–163. https://doi.org/10.1007/s00606-007-0518-1 |
APÉNDICE 1. Características morfológicas, reproductivas, fenológicas, corológicas y altitudinales que diferencian las poblaciones
de Taraxacum aginnense Hofstra, T. ciliare Soest, T. litophyllum De Langhe & Soest, T. raii (Gouan) S.F.Gray y T. gr. mediterraneum
en la península ibérica. [A, ancho; L, largo; + máxima altitud indicada en Granada por Kirschner & Štěpánek (1998).]TOP
| Características |
T. aginnense |
T. ciliare |
T. litophyllum |
T. raii |
T. gr. mediterraneum |
| Contorno de las hojas |
de pinnatífidas a pinnatisectas, raramente dentadas |
dentadas o de pinnatífidas a pinnatisectas |
enteras o ligeramente dentadas |
ligeramente dentadas o de pinnatífidas a pinnatisectas |
de pinnatífidas a pinnatisectas, raramente dentadas |
| L/A del segmento terminal de las hojas |
0,7–1,5 |
(0,6)1,0–2,2(4,4) (a veces ausente) |
ausente |
0,9–2,2 |
(0,9)1,1–1,7 |
| Pecíolo |
Áptero, raramente alado |
Áptero |
alado |
Áptero |
Áptero |
| Disposición de las brácteas externas del capítulo |
adpresas, erectas o raramente patentes |
adpresas o erectas |
adpresas |
adpresas o erectas |
erectas, patentes o ligeramente recurvadas |
| Forma más frecuente de las brácteas externas del capítulo |
ovado-lanceolada |
ovada |
ovado-lanceolada |
ovado-lanceolada |
de lanceolada a ovado-lanceolada |
| L/A de las brácteas externas del capítulo |
(1,3)1,7–2,2(2,5) |
1,1–2,2(2,3) |
3,2–4,3 |
1,4–2,4 |
(1,9)2,2–2,9(3,4) |
| Color de las corolas |
amarillo pálido a amarillo |
amarillo intenso |
amarillo azafranado |
amarillo |
amarillo |
| Tamaño del polen |
irregular |
claramente irregular o rara vez ausente |
ausente |
± regular |
irregular |
| Comportamiento reproductivo |
agamospermia |
agamospermia |
agamospermia |
sexual |
agamospermia |
| L del cono de los aquenios (mm) |
(0,3)0,4–0,6(0,7) |
(0,6)0,8–1,1 |
aquenios inmaduros |
(0,4)0,5–0,8(0,9) |
0,4–0,8 |
| Color de los vilanos |
blanco |
blanco |
blanco, verdoso en la base |
blanco |
blanco |
| Floración (meses) |
III-V |
III-VI |
VII |
III-V |
III-IV |
| Distribución en la península ibérica |
zona litoral del NE de Cataluña |
amplia distribución |
Pirineos centrales |
zona litoral del NE de Cataluña |
zona litoral del NE de Cataluña |
| Rango altitudinal (m) |
0–200 |
0–1650(1900)+ |
c. 1200 |
0–200 |
0–600 |