Revisión taxonómica del género sudamericano Mulinum (Azorelloideae, Apiaceae)
Martina Fernández*, Cecilia Ezcurra & Carolina I. Calviño
Instituto de Investigaciones en Biodiversidad y Medioambiente, CONICET-Universidad Nacional del Comahue, Quintral 1250, 8400 San Carlos de Bariloche, Río Negro, Argentina; mfernandez@comahue-conicet.gob.ar
* Autor para la correspondencia
| |
Resumen
Fernández, M., Ezcurra, C. & Calviño, C.I. 2017. Revisión taxonómica del género sudamericano Mulinum (Azorelloideae, Apiaceae). Anales Jard. Bot. Madrid 74(1): e048.
El género Mulinum es endémico del sur de Sudamérica, donde es ecológicamente importante por su dominancia en la vegetación de regiones elevadas de los Andes y la estepa patagónica. Presentamos un tratamiento en el que se aceptan 10 especies y 3 variedades nuevas. Se aportan una clave para su reconocimiento, listas de sinónimos, descripciones, observaciones sobre afinidades morfológicas entre las especies, discusión de datos bibliográficos, ilustraciones y mapas de distribución. También se analizan y resuelven problemas nomenclaturales, a la vez que se adjuntan listas de nombres dudosos y nombres excluidos. Se reconocen 6 sinónimos nuevos, que incluyen los nombres M. axilliflorum y M. echinus. Mulinum famatinense es considerado aquí una variedad de M. triacanthum. Las otras dos variedades reconocidas son M. spinosum var. hirsutum y M. spinosum var. zechii. Se ha visto ampliada el área de distribución de M. crassifolium, M. leptacanthum, M. spinosum, M. triacanthum y M. ulicinum. Además, se designan 27 lectotipos y un neotipo.
Palabras clave: Andes, estepa patagónica, morfología, nomenclatura, planta en cojín, Umbelliferae.
|
| |
Abstract
Fernández, M., Ezcurra, C. & Calviño, C.I. 2017. Taxonomic revision of the South American genus Mulinum (Azorelloideae, Apiaceae). Anales Jard. Bot. Madrid 74(1): e048.
The genus Mulinum is endemic to southern South America and is ecologically important because of its dominance in the vegetation of the high Andes and the Patagonian steppe. We present a treatment in which 10 species and 3 new varieties are accepted. A diagnostic key to the species is included, and lists of synonyms, morphological descriptions, observations on the morphological affinities between species, review of the relevant literature, illustrations, and distribution maps are provided. We also analyze and resolve nomenclatural problems, and we provide a list of dubious names and excluded names. Six new synonyms are recognized, including the names M. axilliflorum and M. echinus. Mulinum famatinense is considered here a variety of M. triacanthum. The other two varieties recognized are M. spinosum var. hirsutum and M. spinosum var. zechii. The area of distribution for M. crassifolium, M. leptacanthum, M. spinosum, M. triacanthum, and M. ulicinum has been extended. In addition, 27 lectotypes and one neotype are designated.
Keywords: Andes, cushion plant, morphology, nomenclature, Patagonian steppe, Umbelliferae.
|
INTRODUCCIÓNTOP
Mulinum Pers. es un género endémico de Sudamérica y su distribución se restringe a los Andes australes, el centro y sur de los Andes centrales y la Patagonia, donde muchas veces es dominante en los ambientes abiertos donde habita. La mayoría de sus especies son plantas xerófilas, subarbustivas, que forman cojines hemisféricos o chatos, laxos o compactos y crecen en terrenos arenosos, roquedales y pedregales áridos, tanto en la estepa como en la alta montaña.
Hasta el momento, los trabajos acerca de Mulinum han distado de clarificar las complejas relaciones que hay entre varias de las especies del género. Solo contamos con estudios florísticos o catálogos, como los de Clos (1848-1849) y Reiche (1899) para Chile y, para Argentina, el de Pontiroli (1965) para la provincia de Buenos Aires, el de Constance (1988) para la Patagonia y el de Martínez (2003, 2008) para la provincia de San Juan y el Cono Sur —Argentina, sur de Brasil, Chile, Paraguay y Uruguay—. Zech (1992) realizó una revisión taxonómica del género como trabajo de tesis doctoral, sin embargo esta no es considerada una publicación efectiva a efectos nomenclaturales según los artículos n.º 29 y n.º 30 del Código Internacional de Nomenclatura Botánica, ya que nunca fue publicada (McNeill & al., 2012). Este autor dividió el género en secciones, subsecciones y series diferenciadas principalmente por el hábito. Reconoció 8 especies y 8 variedades con énfasis especial en los caracteres diagnósticos asociados a las características de las hojas, la inflorescencia y el fruto, y en la distribución geográfica. Sin embargo, del tratamiento de Zech (1992) queda de manifiesto, a través de las observaciones, que los límites entre varias especies no quedaron resueltos y que las claves propuestas no funcionaban en la práctica para identificarlas.
El último tratamiento del género fue realizado por Martínez (2008). En su listado reconoció 13 especies y mantuvo los nombres y las sinonimias que fueron propuestos por Constance (1988), no así los propuestos por Zech (1992). Por lo expuesto, parecía clara la necesidad de un nuevo estudio del género que resolviera sus problemas taxonómicos y nomenclaturales y, muy especialmente, los límites entre varias de sus especies. Por lo tanto, el objetivo principal de este trabajo fue realizar la revisión taxonómica de Mulinum mediante el análisis de sus colecciones conservadas en herbario y los estudios de campo. En total, en este trabajo se reconocen 10 especies y 3 variedades de Mulinum, se analiza y describe la variación morfológica observada en el género y se discute su historia taxonómica y su afinidad morfológica con otros géneros. Además, se aporta una clave para el reconocimiento de las especies y las variedades aceptadas, listas de sinónimos, descripciones morfológicas, observaciones sobre afinidades morfológicas interespecíficas, discusiones sobre los datos bibliográficos, ilustraciones y mapas de distribución. Se analizan y resuelven problemas nomenclaturales y se añaden listas de nombres dudosos o excluidos.
MATERIAL Y MÉTODOSTOP
Se han estudiado aproximadamente 800 pliegos depositados en los herbarios BA, BAA, BAB, BCRU, BM, CONC, CORD, E, F, G, GOET, K, LAU, LIL, LP, LPB, LY, MA, MCNS, MERL, MPU, NY, P, PR, PRC, SGO, SI, UC y UPS [abreviaturas según Thiers (2015)]. El Apéndice 1 incluye un índice de recolectores y sus números de recolección, con las especies a las que corresponden.
La determinación de las especies y sus sinónimos nomenclaturales y taxonómicos se han realizado tras la comparación del material tipo o las fotografías del mismo y los protólogos correspondientes. En el tratamiento taxonómico, las especies siguen el orden alfabético para facilitar su ubicación en el texto. La cita de los tipos refiere exactamente lo que se lee en las etiquetas de herbario del pliego correspondiente. Entre corchetes se detalla la información que no figura en las etiquetas y que proviene del análisis conjunto de etiquetas y protólogo. La designación de lectotipos se realizó a partir del análisis de los ejemplares o las fotografías de todos los sintipos. Se consultaron las bases de datos disponibles en línea de varios herbarios nacionales y extranjeros, así como la del cibersitio Plant Science en JSTOR (http://plants.jstor.org/). Estas colecciones fotográficas disponibles en JSTOR se designan con el número del herbario o con el número del código de barras seguido de un signo de admiración (!) que indica que dicha imagen ha sido vista. A aquellas colecciones fotográficas no disponibles en JSTOR y que se solicitaron al herbario correspondiente se les añade la abreviatura del herbario donde está depositado el tipo seguida por la expresión “foto!”. Los nombres de los autores se citan de acuerdo con Brummitt & Powell (1992).
Los caracteres morfológicos y las medidas morfométricas utilizados para realizar las descripciones de las especies fueron obtenidos a partir del material de herbario o de ejemplares recolectados durante viajes al campo. Para cada variable asociada a carácter se detalla el rango entre los percentiles del 25% al 75% y, entre paréntesis, se indican los valores de los percentiles menores al 10% o mayores al 90%; estos valores son considerados raros. Para aquellos caracteres cuantitativos que resultaron de utilidad en la clave dicotómica se muestran los gráficos de cajas (box plots) en el Apéndice 2. Se siguió el criterio del Systematics Association Committee for Descriptive Biological Terminology (1962) para la descripción de la forma de las hoja, que incluye lámina, pecíolo y vaina, los segmentos foliares y los frutos. La longitud de los pedicelos florales fue tomada en flores perfectas, ya que no se observó variación interespecífica en el pedicelo de las flores estaminadas. Los mapas de distribución geográfica para cada una de las especies se realizaron a partir de los datos de las etiquetas de herbario de cada ejemplar por medio del programa DIVA-GIS 7.5 (Hijmans & al., 2009).
TRATAMIENTO TAXONÓMICOTOP
Mulinum Pers., Syn. Pl. 1: 309 (1805). TIPO: M. spinosum (Cav.) Pers., designado por Hiroe (1979: 1728).
Azorellopsis H. Wolff, Repert. Spec. Nov. Regni Veg. 19: 312 (1924), syn. nov. TIPO: Azorellopsis trisecta H. Wolff.
Subarbustos olorosos, andromonoicos o androdioicos, ramosos, intrincados, espinescentes o no, que forman cojines hemisféricos o chatos, laxos o compactos, o bien matas bajas y laxas, con ramas viejas densamente cubiertas por restos foliares persistentes. Hojas imbricadas, subsésiles o pecioladas, de rígidas a coriáceas, a veces crasas, trisectas —raro tripartidas—; segmentos planos o cilíndricos, todos con ápice de agudo a muy obtuso o redondeado, punzante o no, glabros o con pelos finos, cortos o largos, con margen entero o, a veces, el de los segmentos laterales de bipartido hasta trífido, glabro —raro con pelos—; base foliar envainadora, la vaina abierta, membranácea, escariosa, con margen ciliado —raro sin cilios—. Inflorescencia en umbelas simples, terminales, situadas desde muy por debajo hasta bien por encima del nivel de las hojas, involucradas, pedunculadas, con 1-34 flores; brácteas involucrales adpresas o reflexas, connadas casi completamente o solo en la base, en forma de cúpula, glabra o densamente lanosa en el interior, con margen entero —raro bi o trilobado y ciliado—. Flores pediceladas; sépalos reducidos a dientes pequeños, triangulares, inconspicuos, persistentes a la madurez; corola blanco-amarillenta, pétalos de elípticos a anchamente elípticos o de ovados a anchamente ovados o espatulados, reflexos, con ápice débilmente inflexo, caedizos en la madurez; estambres más largos que los estilos aunque no sobrepasan los pétalos, con filamentos subulados y anteras globosas, amarillas; estilos 2, divergentes y persistentes; estilopodio de verdoso a amarillento, de cónico a deprimido; ovario ínfero, bicarpelar, bilocular, con 1 primordio seminal por lóculo. Fruto esquizocarpo, de amarillo a rojo, con contorno de elíptico a muy anchamente elíptico o de anchamente obovado a muy anchamente obovado, glabro, con 2 mericarpios comprimidos dorsalmente, con 2 alas por mericarpio, las de ambos mericarpios paralelas o divergentes, con endocarpio leñoso; carpóforo presente, libre o persistentemente unido a uno de los mericarpios, cuando libre entero o bífido.
Historia taxonómica y delimitación del géneroTOP
El género Mulinum fue publicado por primera vez por Persoon (1805) con 4 especies anteriormente incluidas por Cavanilles (1799) en Selinum L. debido a sus umbelas simples y frutos alados y comprimidos dorsalmente. Persoon consideró que estas especies presentaban un hábito distinto al del resto de las especies de Selinum, por lo que debían constituir un género nuevo. Posteriormente, De Candolle (1830) amplió la circunscripción del género y reunió en Mulinum todas las especies de umbelíferas con frutos alados y comprimidos dorsalmente. Dicho autor describió 4 especies nuevas y dividió el género en 3 secciones sobre la base de caracteres vegetativos. Esta concepción del género, sin embargo, fue considerada sumamente artificial y autores posteriores (v.gr., Gillies & Hooker, 1830; Clos, 1848-1849; Weddell, 1857; Philippi, 1860, 1864, 1893-1894; Bentham, 1867; Grisebach, 1874; Baillon, 1880; Echegaray, 1881; Kuntze, 1891-1893; Drude, 1897-1898; Reiche, 1899; Spegazzini, 1899; Macloskie, 1905; Skottsberg, 1916; Wolff, 1921; Constance, 1988; Zech, 1992; Martínez, 2003, 2008) describieron especies nuevas o incluyeron otras especies en el género, a la vez que restringieron el concepto de Mulinum a la “sección I” de De Candolle (1830). Esta sección comprende plantas con frutos alados y comprimidos dorsalmente, de hábito sufruticoso, con hojas 3-5 veces partidas y segmentos a veces espinosos. Esta definición y circunscripción de Mulinum es la que ha permanecido estable por casi dos siglos. Ciertos estudios filogenéticos moleculares (v.gr., Nicolas & Plunkett, 2009, 2012) estimaron que Mulinum así definido no sería monofilético. Esto ya había sido sugerido por numerosos autores debido a la similitud vegetativa entre Mulinum, Azorella Lam. y Laretia Gillies & Hook. (v.gr., Gillies & Hooker, 1830; Weddell, 1857; Bentham, 1867; Baillon, 1880; Constance, 1988; Martínez, 1989). Sin embargo, hasta tanto se realicen estudios filogenéticos y morfológicos comparativos para los tres géneros, mantenemos la circunscripción tradicional de los mismos, establecida sobre la base de los caracteres del fruto.
Todas las especies de Mulinum son subarbustos, es decir, cojines o matas leñosas que no superan los 50 cm de altura. En M. spinosum, esta altura puede superarse con subarbustos hasta de 1 m de altura. La forma de crecimiento en cojín es la más común entre las especies de Mulinum; estas plantas presentan un tallo principal poco desarrollado y muy ramificado desde la base y, según el crecimiento sea uniforme o si prevalece el crecimiento lateral, el contorno de la planta es hemisférico o achatado (Rauh, 1939; Aubert & al., 2014). Por otra parte, el grado de ramificación y el tamaño de los entrenudos y las hojas darán lugar a cojines compactos o laxos. Así, por ejemplo, en M. albovaginatum y M. crassifolium, se observa en un mismo cojín que, en la parte central, la ramificación es más densa, los entrenudos son más cortos y las hojas bien pequeñas en comparación con la periferia, donde la ramificación es más laxa, los entrenudos son más largos y las hojas más grandes. Esto está correlacionado con un crecimiento lento en el centro del cojín y más rápido en la periferia, como ocurre en algunas especies de Azorella (Martínez, 1989). Especies como M. crassifolium o M. spinosum forman cojines hemisféricos y laxos (Fig. 1a). Otras, como M. albovaginatum, M. hallei, M. leptacanthum, M. triacanthum var. famatinense y M. valentini forman cojines chatos, de laxos a compactos, que no superan los 15 cm de altura (Fig. 1b). También es común encontrar matas bajas cuyo crecimiento es irregular y laxo, por lo que no llegan a constituir verdaderos cojines (Fig. 1c); M. echegarayi, M. microphyllum, M. triacanthum var. triacanthum y M. ulicinum presentan este tipo de crecimiento.
|
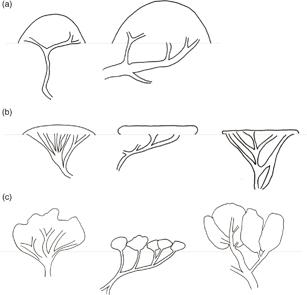 Fig. 1. Esquemas de las distintas formas de crecimiento en Mulinum: a, cojines hemisféricos y laxos de 5-100 cm de altura y 5-300 cm de diámetro (M. crassifolium y M. spinosum); b, cojines chatos, de laxos a compactos, de 2-15 cm de altura y 2-33 cm de diámetro (M. albovaginatum, M. hallei, M. leptacanthum, M. triacanthum var. famatinense y M. valentini); c, matas bajas, irregulares y laxas, de 5-50 cm de altura y 2-30 cm de diámetro (M. echegarayi, M. microphyllum, M. triacanthum var. triacanthum y M. ulicinum). Fig. 1. Esquemas de las distintas formas de crecimiento en Mulinum: a, cojines hemisféricos y laxos de 5-100 cm de altura y 5-300 cm de diámetro (M. crassifolium y M. spinosum); b, cojines chatos, de laxos a compactos, de 2-15 cm de altura y 2-33 cm de diámetro (M. albovaginatum, M. hallei, M. leptacanthum, M. triacanthum var. famatinense y M. valentini); c, matas bajas, irregulares y laxas, de 5-50 cm de altura y 2-30 cm de diámetro (M. echegarayi, M. microphyllum, M. triacanthum var. triacanthum y M. ulicinum).
|
|
En todas las especies de Mulinum las ramas crecen intrincadas, profusamente ramificadas y están cubiertas de restos foliares. La ramificación es simpodial y el crecimiento de cada rama concluye con la formación de una umbela terminal. Las nuevas ramas se originan a partir de las yemas axilares de hojas distales, inmediatamente por debajo de la inflorescencia, como se describió para M. spinosum (Damascos & al., 2008). A veces, los cojines acumulan tierra y humus entre las ramas y alteran, así, las condiciones de temperatura, humedad y concentración de nutrientes en relación con el hábitat que los rodea, lo que facilita el establecimiento y la supervivencia de otras especies en su interior. Este papel de nodriza se ha estudiado en M. leptacanthum (Nuñez & al., 1999; Badano, 2006).
IndumentoTOP
La base foliar de todas las especies de Mulinum es siempre más o menos ciliada, por lo que ninguna de las especies es por completo glabra. En algunas especies, el indumento también está presente en la lámina foliar, en la inflorescencia o en ambas. El indumento se restringe a pelos finos de longitud variable o a cilios —pelos finos y siempre largos, marginales o apicales—, según la especie. Anatómicamente, estos pelos son emergencias cilíndricas, multicelulares, lisas o semiplumosas, con ápice redondeado o truncado (obs. pers.). En este tratamiento, nos referimos a este tipo de indumento como pubescencia en general, sin tener en cuenta en la definición la densidad o la longitud de los pelos, a diferencia de como lo propone, entre otros glosarios, el The Kew Plant Glossary (Beentje, 2010). La presencia de pubescencia no siempre es un carácter constante en todas las hojas de un mismo individuo o de una misma especie. Mulinum albovaginatum es frecuentemente casi glabro, aunque pueda tener pelos cortos y finos esparcidos irregularmente en el margen y la cara adaxial de la lámina foliar. La mayoría de los individuos de M. spinosum también son casi glabros; sin embargo, hemos observado que en algunas poblaciones chilenas el tipo de indumento varía. Así, las poblaciones de Huasco (III Región de Atacama) tienen abundantes pelos largos y finos en toda la hoja, lo que confiere una coloración grisácea característica, mientras que algunas poblaciones de Colchagua (VI Región del Libertador General Bernardo O’Higgins) se distinguen por tener unos pocos pelos muy largos, finos y distribuidos irregularmente por toda la hoja. En M. hallei y M. microphyllum el ápice de los segmentos foliares termina en un cilio fino, largo, blanco y, a veces, caedizo, por lo que suele no verse; la articulación de la lámina con el pecíolo también puede tener cilios. Estas dos especies son las únicas del género que presentan indumento en la inflorescencia. En ellas, la base de las brácteas involucrales tiene cilios extremadamente largos y rizados, que también se encuentran en la base de los pedicelos florales, a los que confiere un aspecto muy lanoso. Asimismo, el pedúnculo de la inflorescencia y el margen y el ápice de las brácteas pueden tener cilios similares a los que se encuentran en las hojas. Los frutos de Mulinum son siempre glabros.
Morfología foliarTOP
Las hojas de Mulinum están divididas en 3 segmentos, generalmente son rígidas, punzantes, pecioladas y tienen una base envainadora. Si bien en este sentido son bastante homogéneas, su tamaño y la consistencia y la forma de los segmentos y el ápice son caracteres sumamente útiles para diferenciar las especies.
El tamaño de las hojas —lámina, pecíolo y vaina— varía de los 4 mm de longitud en M. albovaginatum o M. valentini, a los 8 cm en M. spinosum. La consistencia de las hojas es, en la mayoría de las especies, rígida, aunque en M. hallei y M. microphyllum es coriácea y en M. crassifolium y M. echegarayi, crasa. Las hojas huelen cuando se estrujan o rompen debido a que tienen canales oleíferos. Las láminas foliares son trisectas o, menos frecuentemente, tripartidas. A veces, los segmentos laterales pueden ser bipartidos y puede resultar una lámina con 5 segmentos aparentes (v.gr., en M. hallei, M. leptacanthum, M. microphyllum y M. spinosum). En M. microphyllum los segmentos laterales también pueden ser tripartidos, por lo que las láminas pueden tener hasta 7 segmentos aparentes. Los segmentos pueden ser planos o cilíndricos y de forma y anchura variables. El ápice puede ser agudo o de obtuso a redondeado, mucronado —v.gr., en M. crassifolium, M. echegarayi, M. hallei y M. microphyllum—, espinuloso —M. albovaginatum y M. valentini— o espinoso —en el resto de las especies—. El pecíolo es similar a los segmentos. Las hojas pueden ser casi sésiles —muy cortamente pecioladas; v.gr., en M. echegarayi y M. valentini— o pecioladas —en el resto—. En M. albovaginatum y M. crassifolium las hojas que se encuentran en el centro del cojín son menos pecioladas que aquellas de la periferia. La base foliar es membranácea, generalmente amplexicaule y forma una vaina abierta. Sin embargo, en algunos ejemplares de M. leptacanthum y M. spinosum la base foliar está poco ensanchada, por lo que no rodea completamente el tallo.
InflorescenciaTOP
La inflorescencia en Mulinum se compone de una umbela simple, terminal, con un número variable de flores y brácteas. Las umbelas son pedunculadas y pueden estar muy por debajo del nivel de las hojas del cojín —v.gr., en M. triacanthum y M. ulicinum—, apenas sobrepasar este nivel —v.gr., en M. albovaginatum, M. crassifolium, M. echegarayi, M. leptacanthum y M. valentini—, o bien sobrepasarlo ampliamente —v.gr., en M. hallei y M. microphyllum—. En M. spinosum hay de umbelas que apenas sobrepasan el nivel de las hojas a otras que lo sobrepasan ampliamente. En M. triacanthum y M. ulicinum se observa la mayor reducción de los pedúnculos y pedicelos en relación al tamaño de las hojas.
Las brácteas del involucro son más cortas que los pedicelos florales y poco conspicuas. En la mayoría de las especies son adpresas y pueden estar connadas hasta la mitad o solo en la base, en forma de una cúpula más o menos profunda. En M. spinosum las brácteas del involucro pueden ser reflexas. En M. hallei y M. microphyllum las brácteas involucrales son siempre reflexas, están connadas solo en la base y forman una cúpula densamente lanosa en su interior, como se ha comentado previamente.
Las umbelas son generalmente homogéneas, es decir, que todas sus flores son perfectas o estaminadas. Las especies pueden ser andromonoicas o androdioicas. En M. spinosum también hay umbelas heterogéneas con flores estaminadas en la periferia y perfectas en la parte central. A lo largo del desarrollo de la umbela, van ocurriendo distintos estadios fenológicos hasta la maduración y escisión de los mericarpios, que caen por acción de la gravedad o el viento. El desarrollo de las umbelas —como ocurre en la mayoría de las apiáceas (Webb, 1981) y se describió para Azorella (Martínez, 1989)— es abierto y la maduración es centrípeta; así, los botones florales van madurando y fructificando desde la periferia hacia el interior de la umbela a medida que se van formando.
En Mulinum las flores son pequeñas y pediceladas. Los sépalos son persistentes y están reducidos a pequeños dientes triangulares, mucho más cortos que los pétalos. Los pétalos confieren el color amarillo a las flores y son, en la mayoría de las especies, de elípticos a ovados y de anchura variable, o bien espatulados y marcadamente angostos en la base —M. hallei y M. microphyllum—; en todos los casos son reflexos, tienen el ápice débilmente inflexo y se tornan caedizos cuando maduran. Los estambres son más largos que los estilos pero más cortos que los pétalos y tienen filamentos subulados y anteras globosas y amarillas. Los estilos son 2, divergentes y persistentes; el estilopodio es, en la mayoría de las especies de Mulinum, un disco de cónico a deprimido que, en todos los casos, es de menor diámetro que la longitud de los estilos.
Todas las especies de Mulinum tienen individuos con flores estaminadas, perfectas o de ambos tipos. La proporción de cada tipo de flor es variable en cada especie. No se han observado individuos de especie alguna con flores pistiladas, a pesar de que Constance (1988) mencionó esta característica para M. hallei, M. leptacanthum, M. spinosum y M. valentini.
Mulinum tiene el fruto característico de la familia Apiaceae; esto es, un esquizocarpo que se separa en dos mericarpios. Cada mericarpio está comprimido dorsalmente y es alado. Los frutos son glabros, de amarillos a rojos, de tamaño variable y contorno de elíptico a circular. Los dos mericarpios están unidos por sus comisuras y, en la madurez, se separan y dejan ver el carpóforo. En Mulinum, el carpóforo es generalmente libre —se separa de ambos mericarpios fácilmente—, como ocurre en M. echegarayi, M. hallei, M. microphyllum, M. triacanthum y M. ulicinum. Sin embargo, en M. albovaginatum, M. crassifolium y M. valentini, el carpóforo no es completamente libre, ya que se separa solo de uno de los mericarpios en la madurez y persiste unido al otro —puede separarse con ayuda mecánica de este mericarpio, pero con dificultad—. En M. leptacanthum y M. spinosum, el carpóforo puede ser completamente libre o no. Cuando el carpóforo es libre, el grado de división del ápice varía en todas las especies de entero a cortamente bífido —se divide hasta la base solo en M. spinosum.
Cada mericarpio está recorrido longitudinalmente por 5 costillas bien marcadas: una dorsal sobre el eje central de la superficie dorsal del mericarpio, 2 laterales prolongadas en alas y otras 2 marginales o comisurales en la superficie comisural del mericarpio. La relación entre la anchura de las alas y la del cuerpo del fruto varía entre especies. Así, las alas pueden tener la misma anchura o ser más anchas que el cuerpo del fruto —v.gr., en M. crassifolium, M. echegarayi, M. hallei, M. leptacanthum, M. microphyllum, M. spinosum, M. triacanthum var. triacanthum y M. valentini— o ser muy angostas y configurar un fruto casi áptero —v.gr., en M. triacanthum var. famatinense y M. ulicinum—. En ciertos trabajos que incluyen a Mulinum (cf. Tseng, 1967; Constance, 1988), se habla de frutos con 2 o 4 alas; estrictamente todos los frutos tienen 4 alas (2 por mericarpio), sin embargo, cuando las alas de cada mericarpio están separadas 180° y se alinean sobre el plano comisural, parece que hay solo 2 alas por fruto, ya que las alas de mericarpios distintos quedan en paralelo y en contacto. En cambio, cuando las alas de cada mericarpio divergen y están separadas por menos de 180°, las 4 alas se distinguen claramente. En este trabajo, hemos decidido reemplazar la anterior nomenclatura que hablaba de frutos con 2 o 4 alas por frutos con alas paralelas o divergentes, respectivamente, ya que la consideramos más apropiada y comprensible. En Mulinum es más común encontrar frutos con alas divergentes, aunque con diferentes grados de apertura de las alas. Sin embargo, el grado de apertura es variable incluso dentro de una misma especie, por lo que no se le ha encontrado valor taxonómico alguno, a excepción de la distinción extrema entre alas paralelas o divergentes.
Distribución geográfica y hábitatTOP
Mulinum es un género endémico de Sudamérica y característico de regiones elevadas de los Andes de Chile, el sur de Bolivia y Argentina y de la estepa patagónica. Se extiende aproximadamente a partir de los 17º de latitud norte hasta los 53º de latitud sur y desde el nivel del mar hasta los 4900 m s.n.m. En Argentina, el género está presente desde el noroeste del país —provincia de Jujuy— hasta el extremo sur continental —provincia de Santa Cruz—; en Chile, desde la I Región de Tarapacá hasta la parte continental de la XII Región de Magallanes y de la Antártica Chilena; y en Bolivia la distribución de Mulinum se restringe al sur, en los departamentos de Cochabamba, Oruro y Tarija. La mayoría de las especies crecen en terrenos arenosos, roquedales y pedregales áridos del dominio Andino-Patagónico en las provincias fitogeográficas Puneña, Altoandina y Patagónica (Cabrera, 1971; Cabrera & Willink, 1980).
Las 10 especies del género habitan en Argentina, 6 de ellas también en Chile (Fig. 2) —v.gr., M. albovaginatum, M. crassifolium, M. leptacanthum, M. spinosum, M. ulicinum y M. valentini— y una sola también en Bolivia, M. triacanthum. Las 3 especies restantes, M. echegarayi, M. hallei y M. microphyllum, son endémicas de Argentina. La especie con distribución más restringida es M. echegarayi, endémica de las cordilleras de San Juan y Mendoza. Por el contario, M. spinosum es la especie más ampliamente distribuida y la única que presenta áreas de distribución disyuntas, ya que además de vivir a lo largo de los Andes y la Patagonia —desde San Juan hasta Santa Cruz en el oeste y el sur de Argentina y las regiones limítrofes de Chile—, también lo hace en la región pampeana, en las Sierras Pampeanas —provincia de San Luis— y en la Sierra de la Ventana —provincia de Buenos Aires—, en el centro de Argentina.
|
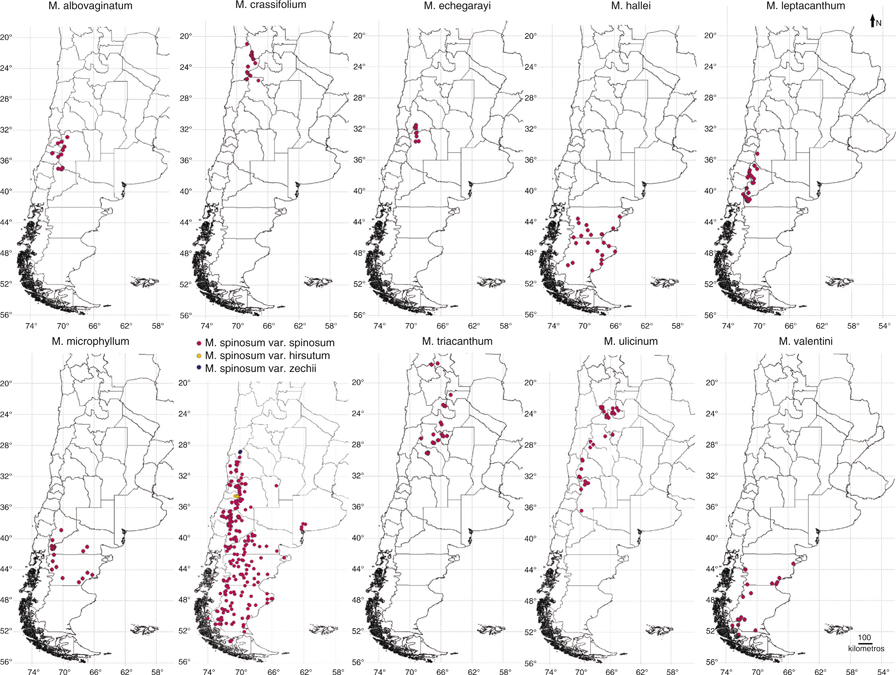 Fig. 2. Mapas de distribución de las especies de Mulinum. Fig. 2. Mapas de distribución de las especies de Mulinum.
|
|
Clave para determinar las especies de MulinumTOP
| 1. |
Ápice de los segmentos foliares no punzante y terminado en un cilio fino, largo y blanco, caedizo; brácteas del involucro en forma de cúpula densamente lanosa en el interior; pétalos espatulados, con la base marcadamente angosta
|
2 |
| – |
Ápice de los segmentos foliares punzante y mucronado, espinuloso o espinoso; brácteas del involucro en forma de cúpula glabra en el interior; pétalos ni espatulados, ni con la base angosta
|
3 |
| 2. |
Cojines chatos y compactos; segmentos foliares no articulados en su inserción, a veces, algunos del mismo individuo articulados; hojas (5)7-10(11) mm; lámina foliar de (2)3-4(6) mm de anchura; pecíolo 1-3(4) mm; pedúnculos (6)8-11(16) mm
|
4. M. hallei |
| – |
Matas bajas y laxas; segmentos foliares articulados en su inserción; hojas (9)11-14(15) mm; lámina foliar de 6-8(9) mm de anchura; pecíolo (2)3-6(7) mm; pedúnculos (11)19-30(39) mm
|
6. M. microphyllum |
| 3. |
Segmento foliar central 1-4 mm, siempre plano, con una relación longitud/anchura de menos de 2(2,5), con ápice mucronado o espinuloso
|
4 |
| – |
Segmento foliar central (3,5)4-19(23) mm, cilíndrico o plano, con una relación longitud/anchura de (1,5)2 o más, con ápice espinoso
|
7 |
| 4. |
Carpóforo libre; matas bajas y laxas hasta de 50 cm de altura y 20 cm de diámetro; plantas androdioicas, con pedicelos florales de 3-4,5 mm
|
3. M. echegarayi |
| – |
Carpóforo persistentemente unido a uno de los mericarpios; cojines hemisféricos y laxos, o bien chatos y compactos, hasta de 20 cm de altura y 40 cm de diámetro; plantas andromonoicas, si androdioicas, entonces con pedicelos florales de 2-3 mm
|
5 |
| 5. |
Cojines androdioicos, hemisféricos y laxos; hojas crasas; segmentos foliares de anchamente a muy anchamente ovados, con una relación longitud/anchura de menos de 1,4; distribución en Salta, Argentina, y en las regiones I-III, Chile
|
2. M. crassifolium |
| – |
Cojines andromonoicos, chatos y compactos; hojas no crasas; segmentos foliares de angostamente ovados a ovados, con una relación longitud/anchura de más de 1,5, o bien de triangulares a anchamente triangulares; distribución en Argentina desde Mendoza al sur, y en Chile en regiones limítrofes
|
6 |
| 6. |
Hojas (4)6-8(10) × (2)3-4(9) mm, glabras o, a veces, pubescentes en la cara adaxial; segmentos foliares de angostamente ovados a ovados; fruto de (2)3-4(5) mm de anchura, con alas de 0,5-1 mm de anchura, de la misma anchura o más angostas que el cuerpo del fruto; distribución en Mendoza y Neuquén, Argentina, y en las regiones RM y VI, Chile
|
1. M. albovaginatum |
| – |
Hojas 8-13(15) × 4-6(7) mm, siempre glabras; segmentos foliares de triangulares a anchamente triangulares; fruto de (3)5-6(7) mm de anchura, con alas de 1-3 mm de anchura, más anchas que el cuerpo del fruto; distribución en Chubut y Santa Cruz, Argentina y en la región XII, Chile
|
10. M. valentini |
| 7. |
Pedúnculos 11-43(50) mm; cojines hemisféricos y laxos; hojas (9)16-43(51) mm; umbelas que apenas sobrepasan o que sobrepasan netamente el nivel de las hojas
|
7. M. spinosum |
| – |
Pedúnculos (2)3-6(10) mm; cojines chatos y compactos o matas bajas, irregulares y laxas; hojas de menos de 17(20) mm, o si más largas, entonces con umbelas muy por debajo del nivel de las hojas
|
8 |
| 8. |
Segmentos foliares planos; cojines chatos y compactos con frutos de más de 5 mm de anchura. |
5. M. leptacanthum |
| – |
Segmentos foliares cilíndricos; matas bajas y laxas con frutos generalmente de menos de 5 mm de anchura, o si cojines chatos y compactos, entonces frutos de menos de 4 mm de anchura
|
9 |
| 9. |
Cojines chatos y compactos con frutos muy comprimidos dorsalmente, o bien matas bajas y laxas con el segmento central de la lámina de más de 9 mm
|
8. M. triacanthum |
| – |
Matas bajas y laxas con frutos poco comprimidos dorsalmente y con el segmento central de la lámina de menos de 7 mm |
9. M. ulicinum |
1. M. albovaginatum Gillies & Hook., Bot. Misc. 1: 328-329 (1830). TIPO: [Argentina:] Mendoza, Cordillera, Gillies s.n. (lectotipo, designado aquí: K 000529788 foto!). Fig. 3.
|
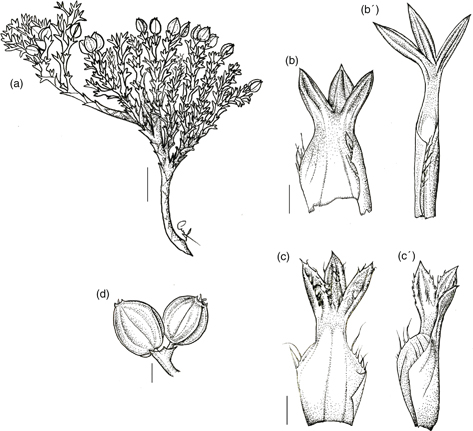 Fig. 3. Mulinum albovaginatum: a, fragmento del cojín; b, b’, hojas del mismo individuo tomadas del centro y de la periferia del cojín; c, c’, hoja pubescente en distintas posiciones; d, umbela con frutos [a, b, b’, d, Boelcke 11228 (SI); c, Boelcke 11610 (SI)]. [Escala: a = 1 cm; b, c = 2 mm; d = 2,5 mm.] Fig. 3. Mulinum albovaginatum: a, fragmento del cojín; b, b’, hojas del mismo individuo tomadas del centro y de la periferia del cojín; c, c’, hoja pubescente en distintas posiciones; d, umbela con frutos [a, b, b’, d, Boelcke 11228 (SI); c, Boelcke 11610 (SI)]. [Escala: a = 1 cm; b, c = 2 mm; d = 2,5 mm.]
|
|
M. pauciflorum Reiche, Anales Univ. Chile 104: 803 (1899), syn. nov. M. reichei H. Wolff, Repert. Spec. Nov. Regni Veg. 17: 441-442 (1921), nom. illeg. M. albovaginatum var. pauciflorum (Reiche) Zech, Kurtziana 24: 192 (1995). TIPO: [Chile, VII Región del Maule:] Cordillera de Curicó, 2500 m, Reiche s.n. (lectotipo, designado aquí: SGO 000003755 foto!; isolectotipo: B destruido, foto F neg. 003453!).
M. cryptanthum var. pulvinaris Chodat & Wilcz., Bull. Herb. Boissier 2: 525 (1902). TIPO: [Argentina, Mendoza:] Cajón del Burro, Valle del Río Tordillo, 2900 m, Wilczek 65 (lectotipo, designado aquí: G 00358897 foto!; isolectotipo: G 00358898 foto!).
Subarbusto andromonoico que forma cojines de 2-10 cm de altura y 20-30 cm de diámetro, chatos y compactos. Hojas (4)6-8(10) mm, subsésiles las que se encuentran en el centro del cojín o pecioladas las de la periferia, de obovadas a anchamente obovadas u oblongas, de semirígidas a rígidas; lámina (2)2,5-4(5) × (2)3-4(9) mm, trisecta con el segmento central de (1,5)2-4 × 1-1,8(2) mm; segmentos de angostamente ovados a ovados, planos, surcados en la cara adaxial, glabros o con pelos cortos y finos en la cara adaxial, con ápice obtuso los del centro del cojín o agudo en los de la periferia, espinuloso, punzante, con margen entero, glabro o con pelos esparcidos irregularmente; pecíolo 1-2 mm, más ancho que los segmentos en las hojas subsésiles o semejante a los segmentos en las pecioladas, glabro; base foliar amplexicaule, con margen ciliado, raro sin cilios. Umbelas que apenas sobrepasan el nivel de las hojas, con 1-4 flores perfectas o 5-7 flores estaminadas; pedúnculos 3-4(6) mm, glabros; involucro de 2-4 brácteas de 1,5-3 mm, adpresas, connadas casi completamente en forma de cúpula profunda con lóbulos de triangulares a anchamente triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de (2)3-5 mm, glabros; pétalos 1-2 × 1,2-1,5 mm, de ovados a anchamente ovados; estilopodio de algo cónico a deprimido, los estilos 0,8-1 mm. Fruto (3)5-6 × (2)3-4(5) mm, de amarillento a rojizo, de contorno circular a anchamente elíptico, poco comprimido dorsalmente; mericarpio de 1-2 mm de espesor; alas de 0,5-1 mm de anchura, de la misma anchura o más angostas que el cuerpo del fruto, divergentes; carpóforo persistentemente unido a uno de los mericarpios.
Distribución geográfica, hábitat y fenología.—Se distribuye por el oeste de Argentina —provincias de Mendoza y Neuquén— y el centro de Chile —Región Metropolitana de Santiago de Chile y VII Maule—, en la Cordillera de los Andes (Fig. 2). Habita en terrenos rocosos; 2000-3800 m s.n.m. Florece de noviembre a febrero y fructifica de diciembre a principios de abril.
Observaciones.—Se amplía la distribución de esta especie a la Región Metropolitana de Santiago de Chile. Por otra parte, Zech (1992) mencionó un único ejemplar de esta especie (Bridges s.n., BM) de la VI Región del Libertador General Bernardo O’Higgins de Chile, que no pudimos localizar; Martínez (2008) hizo mención de esta cita. Sin embargo, entre todo el material de herbario estudiado, recolectado en Chile y depositado en distintos herbarios, no hemos encontrado ejemplar alguno de M. albovaginatum de dicha región, por lo que su presencia allí es dudosa.
Mulinum albovaginatum se asemeja a M. valentini, pero tiene hojas menores —6-8 mm—, segmentos foliares ovados y frutos más angostos —3-4 mm—, mientras que M. valentini tiene hojas de 8-13 mm, segmentos foliares triangulares y frutos de 5-6 mm. Además, la distribución de estas dos especies es alopátrica: M. albovaginatum vive en Mendoza y Neuquén en Argentina y en las regiones Metropolitana de Santiago y VII del Maule de Chile, mientras que M. valentini más al sur, en Chubut y Santa Cruz en Argentina y, en Chile, en la XII Región de Magallanes y de la Antártica Chilena.
Reiche (1899) consideró M. albovaginatum semejante a Azorella spinosa (Ruiz & Pav.) Pers. Sin embargo, ambas especies son fácilmente diferenciables: esta última se caracteriza por tener la lámina foliar rómbica, con venas blanquecinas muy marcadas en la cara adaxial, de mayor tamaño —7-15 mm—, segmentos foliares triangulares y frutos de contorno oblongo y ápteros (Calviño & al., 2016).
Mulinum pauciflorum, aquí tratado como sinónimo de M. albovaginatum, fue considerado una nueva especie por Reiche (1899), por tener la lámina foliar cubierta de pelos blancos y frutos grandes de 8 mm. Zech (1992, 1995), sobre la base de estos mismos caracteres y de la distribución geográfica restringida a Chile —regiones VI, Libertador General Bernardo O’Higgins y VII, Maule—, trató esta entidad como una variedad de M. albovaginatum —M. albovaginatum var. pauciflorum—. Por otra parte, consideró que la variedad albovaginatum, con láminas glabras y menores frutos, se restringe a Argentina —provincias de Mendoza y Neuquén—. Sin embargo, el estudio del material de herbario nos ha permitido determinar que hay ejemplares recolectados en Chile con lámina foliar glabra —v.gr., Mieres s.n., CONC 166141; Teillier 4556, CONC— y ejemplares recolectados en Argentina con lámina foliar pubescente —v.gr., Villagrán & al. 8128, CONC; Boelcke & al. 11610, SI; Castellanos s.n., BA 36808—; además, el tamaño de los frutos varía independientemente de la pubescencia de las hojas y la distribución geográfica. En consecuencia, no hay una asociación entre la pubescencia y el tamaño del fruto, ni entre la variación morfológica de la especie y su distribución, por lo que consideramos que M. albovaginatum var. pauciflorum no tiene sustento y es sinónimo de M. albovaginatum.
Designamos aquí el pliego recolectado por Gillies s.n., K 000529788, como lectotipo de M. albovaginatum entre este y los sintipos E 00000042 y E 00000043, porque el pliego de K tiene material más abundante. Para M. pauciflorum se designa el pliego SGO 000003755 como lectotipo, ya que de su duplicado destruido de B sólo se dispone de una fotografía (F neg. 003453). Para M. cryptanthum var. pulvinaris se selecciona el pliego recolectado por Wilczek 65, G 00358897, poque tiene una mayor cantidad y calidad de material que el resto de material tipo —v.gr., Wilczek 65, G 00358898; Cumming 1188, W 0013100; Bridges 1842, G 00309596.
Material estudiado.—ARGENTINA. Mendoza: Andes, Gillies s.n. (E 00000042). Dpto. Luján de Cuyo, Cerro de la Polcura, Gillies s.n. (E 00000043). Dpto. Malargüe, trayecto de Malargüe a Las Loicas, 1400-1750 m, 29-I-1994, Villagrán & al. 8128 (CONC); Valle de Las Leñas, portezuelo ancho a Valle Encantado, 2650-2800 m, 11-II-1987, Kiesling 6763 (SI); Meseta de La Sierra de la Media Luna, 2380 m, 22-I-1981, Méndez & Willoud 31495 (MERL); inmediaciones del refugio militar Valle del Atuel, 2100 m, 9/17-I-1954, Ruiz Leal & Roig 17563 (MERL); Portezuelo del Choique, 2350 m, 14-XII-1960, Ruiz Leal & al. 21439 (MERL, UC); 07-II-1950, Boelcke & al. 4217 (BAB, UC); proximidades de cresta en Sierra Azul, 2780 m, 12-III-1981, Méndez & Willoud 31516 (MERL); Alto Valle del Atuel, desembocadura del arroyo, 9/17-I-1954, Ruiz Leal & Roig 15654 (MERL, UC); 2-4 km NO de Las Leñas, 3080 m, 7-III-1987, Stuessy & al. 10333 (CONC, LP, UC); La Valenciana, 2100 m, 4-II-1942, Ruiz Leal & al. 7828 (BA); Paso Planchón, 18-I-1941, Castellanos s.n. (BA 36808); Cajón del Burro, Valle del Río Tordillo, 2900 m, Wilczek s.n. (LAU). Dpto. San Carlos, camino a Laguna Diamante, 3000 m, 27-I-1950, Cuezzo & Barkley 20Mz383 (LIL, UC); 2750 m, 2-II-1950, Boelcke & al. 4624 (BAB, UC); Laguna Diamante, 3300 m, 17-I-1989, Zech & Contogiorgakis 9 (LP); 3300 m, 4-III-1943, Covas 10827 (MERL); 10-III-1943, Covas 10828 (MERL); Laguna Diamante, bajada al este de El Paramillo, 18-I-1963, Boelcke & al. 10064 (SI); 28,8 km de RN 98 hacia Lago Diamente por RN 101, 3700 m, 22-I-2002, Chandler & Bayer 1114 (LP); RN 98, Pampa de los Avestruces, 3780 m, 25-I-1985, Hunziker & al. 11310 (SI); arroyo de los Gauchos; 19-I-1965, Roig & Ruiz Leal 45571 (MERL); arroyo de los Gauchos, ribera oeste, 18-I-1963, Boelcke & al. 10033 (SI); Estancia Llaucha, Pampa de las Osamentas, 18-I-1941, Ruiz Leal & al. 7183 (BA, LIL, MERL, UC); inmediaciones de Las Vegas de Llaucha, II-1942, Patiño 7917 (MERL); Quebrada del Paso de la Cruz de Piedra, aduana, 15-I-1949, Ruiz Leal & al. 11682 (MERL, UC); 16-I-1952, Ruiz Leal & al. 14578 (MERL, UC). Dpto. San Rafael, Cerro Nevado, I-1926, Carette 2515 (BA 76816, MERL); Las Ánimas, Río Salado, 12-II-1946, Rossi 317 (LIL); Los Molles, 8/10-XII-1969, Sosa 27132 (MERL); Vega del Loro, 2400 m, -I-1921, Carette 290 (SI); arroyo Negro, cerca de las lagunas, 7/8-I-1888, Kurtz 5756 (CORD); Portezuelo del Calqueque, 28-I-1888, Kurtz 5905a (CORD); Calmuco, 2600 m, 14-II-1942, Covas 10913 (MERL, UC). Dpto. Tunuyán, 20,3 km al O de Manzana Histórica, O de Tunuyán a lo largo del arroyo Grande, 3138 m, 20-I-2002, Chandler & Bayer 1105 (LP); Paso del Portillo, Cuesta de los Afligidos, 27-XII-1933, Ruiz Leal & al. 1983 (BA, MERL); nacimientos del Arroyo de las Cuevas, 3225 m, 20/21-III-1935, Ruiz Leal & al. 3155 (BA, MERL, UC); rincones del cerro Morado, 3700 m, 21-III-1935, Ruiz Leal & al. 3169 (BA, MERL, UC); sierra del Nevado, lomas al SSE expuesta al S, 2700 m, 22-I-1974, Boelcke & al. 15927 (SI). Dpto. Tupungato, Estancia Silva, Loma Larga (mogote Bolvaran), 3000 m, 10-IV-1934, Salinas 2180 (LIL, MERL); Tres Quebradas, 2840 m, 12-XII-1935, Ruiz Leal & al. 3619 (BA, MERL). Neuquén: Dpto. Chos Malal, 25 km al NW de Chos Malal, rumbo a Andacollo, arroyo Chacay-Melehue, 17-XI-1969, Ancibor & al. 90214 (BAB, UC); Parque Provincial Tromen, cerro Waile, cumbre a 3100 m, 29-XII-1999, Ezcurra & al. 2666 (BCRU); a 45 km del cruce de RN 40, 2100 m, 29-XII-1999, Ezcurra & al. 2581 (BCRU); RP 37, 60 km al norte de Chos Malal, ladera exposición oeste, 16-I-1995, Chiapella & al. 544 (BCRU); RP 37, 55 km al norte de Chos Malal, camino a Barrancas, 7-II-1994, Chiapella & al. 284 (BCRU); cajón del arroyo del Cruce, 2500 m, 27-I-1964, Boelcke & al. 11228 (SI, UC). Dpto. Minas, Cordillera del Viento, cruzada de Tricao Malal al Cajón de Butaló, ladera este, 3-XI-1964, Boelcke & al. 11610 y 11613 (BAB, SI, UC); a 33 km de Tricao Malal camino a Mina de Azufre, 2350 m, 5-II-1964, Boelcke & al. 11633 (BAB, SI, UC). Dpto. Zapala, Parque Nacional Laguna Blanca, II-1958, Perrone s.n. (BA 30254).
CHILE. Sine loc., Bridges 1842 (G 00309596); Cumming 1188 (W 0013100). Región Metropolitana de Santiago: Prov. Cordillera, Paso de Maipo, sector Picos Bayos, 3440 m, 23-III-2006, Mieres s.n. (CONC 166141); Cajón del Maipo, Piedras Negras, 3350 m, 20-I-2000, Teillier 4556 (CONC). VII Región del Maule: Pcia. Curicó, Cordillera del volcán Peteroa, Werdermann 613 (SI, UC); Valle del río Teno, 1000 m, 29-XII-1971, Zölner 5950 (CONC, UC).
2. M. crassifolium Phil., Fl. Atacam.: 24 (1860). TIPO: [Chile, II Región de Antofagasta, 23°52’ lat.:] Desierto de Atacama, Alto de Puquios, 4000 m, II-1854, Philippi s.n. (lectotipo, designado aquí: SGO 000003745 foto!). Fig. 4.
|
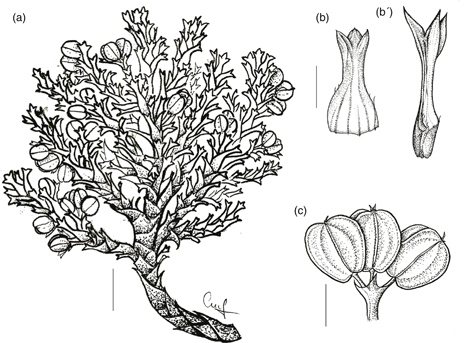 Fig. 4. Mulinum crassifolium: a, fragmento del cojín; b, b’, hojas del mismo individuo tomadas del centro y de la periferia del cojín; c, umbela con frutos [Latorre & al. 206 (CONC)]. [Escala: a = 1 cm; b, b’ = 2 mm; c = 2,5 mm.] Fig. 4. Mulinum crassifolium: a, fragmento del cojín; b, b’, hojas del mismo individuo tomadas del centro y de la periferia del cojín; c, umbela con frutos [Latorre & al. 206 (CONC)]. [Escala: a = 1 cm; b, b’ = 2 mm; c = 2,5 mm.]
|
|
Azorellopsis trisecta H. Wolff, Repert. Spec. Nov. Regni Veg. 19: 312 (1924), syn. nov. TIPO: [Chile, II Región de Antofagasta:] Pcia El Loa, a 70 km de San Pedro de Atacama por RN 5, 3 km al sur del desvío a Bolivia, 4460 m, 14-XII- 2010, Calviño & al. 789 (neotipo, designado aquí: SI!; isoneotipo: BCRU!).
Subarbusto androdioico que forma cojines de 5-20 cm de altura y 15-40 cm de diámetro, hemisféricos, laxos y resinosos. Hojas 7-8 mm, subsésiles las de la parte central del cojín o pecioladas las de la periferia, de ovadas a anchamente ovadas, crasas, rígidas; lámina 2-4 × (2)3,5-4,5(9) mm, trisecta o tripartida, con el segmento central de 1-2,5(3) × (1,6)1,8-2,2 mm; segmentos de anchamente ovados a muy anchamente ovados —raro deprimido-ovados—, planos, surcados en la cara adaxial, glabros, con margen entero y ápice de muy obtuso a redondeado en los de la parte central del cojín o agudo en los de la periferia, mucronado, punzante; pecíolo 1-1,5(2) mm, poco más ancho que los segmentos en las hojas subsésiles o semejante a los segmentos en las pecioladas, glabro; base foliar amplexicaule, margen escasamente ciliado —raro sin cilios—. Umbelas que apenas sobrepasan el nivel de las hojas, con 2-5 flores perfectas o 5-10 flores estaminadas; pedúnculos 2-3,5 mm, glabros; involucro de 2-3 brácteas de 1,5-3 mm, adpresas, connadas casi completamente en forma de cúpula profunda con lóbulos triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 2-3 mm, glabros; pétalos 1,2-2 × 1-1,2 mm, de elípticos a anchamente elípticos; estilopodio deprimido y estilos 1-1,2 mm. Fruto 3-6 × 3-5 mm, amarillento, de contorno de anchamente obovado a muy anchamente obovado, poco comprimido dorsalmente; mericarpio de 1-2 mm de espesor; alas de 1-2 mm de anchura, de la misma anchura o más anchas que el cuerpo del fruto, divergentes; carpóforo persistentemente unido a uno de los mericarpios.
Distribución geográfica, hábitat y fenología.—Se distribuye por el noroeste de Argentina —provincias de Salta y Catamarca— y el norte de Chile —I Tarapacá, II Antofagasta y III Atacama—, en la Cordillera de los Andes (Fig. 2). Habita en abras planas y expuestas, en suelo pedregoso; 3700-4600 m s.n.m. Florece de noviembre a mediados de febrero y fructifica de diciembre a febrero.
Nombres vernáculos y usos.—Zucurco” (Philippi, 1860); chuquicán y sulultur (Munizaga & Gunckel, 1958); sucurco y espinilla (De Mösbach, 1992); chuchikan, chukikandi, chukikandia y chuchikandia (en Chile, Villagrán & al., 1998a,b).
Es una especie muy utilizada y considerada “poderosa” en la medicina tradicional de las comunidades atacameñas de la II Región de Antofagasta en Chile (Villagrán & al., 1998b). Se le atribuyen propiedades digestivas, es remedio contra las afecciones bronquiales, el resfriado, la tos, el dolor de cabeza y la diabetes; también es útil para contrarrestar los efectos del mal de altura o puna (Munizaga & Gunckel, 1958; Niemeyer, 1995; Villagrán & al., 1998a,b).
Observaciones.—Se amplía la distribución de esta especie a la provincia de Catamarca, Argentina, y a la I Región de Tarapacá de Chile, donde. es la única representante del género. En la II Región de Antofagasta vive con M. ulicinum, por lo que se las ha confundido. Sin embargo, estas especies son fácilmente diferenciables por medio de ciertos caracteres relacionados con su hábito, hojas y frutos. Mulinum ulicinum forma matas bajas, las hojas son de mayor tamaño —13-17 mm—, los segmentos foliares son triangulares y de sección circular y su ápice es agudo y espinoso, además, el fruto tiene contorno elíptico y alas más angostas que su cuerpo; M. crassifolium forma cojines hemisféricos, las hojas son de 7-8 mm, tiene segmentos foliares ovados y de sección plana, con ápice de muy obtuso a redondeado y mucronado y el fruto contorno anchamente obovado y alas de la misma anchura o más anchas que su cuerpo (Fernández & al., 2014). La forma hemisférica de los cojines, sus hojas carnosas y una resina de transparente a color ambarino que se encuentra en todas las partes de la planta, confieren a M. crassifolium un aspecto característico.
En 1924, Wolff publicó por primera vez el género Azorellopsis y mencionó en el protólogo que era parecido a Azorella por la forma de crecimiento, pero que se diferenciaban por la estructura del ovario —con costillas dorsales gruesas y las laterales aladas en Azorellopsis—. A la vez, publicó la especie Azorellopsis trisecta y citó un material recolectado por Bridges en Bolivia, sin referencia a localidad ni fecha de colección y depositado en W. En este herbario no se ha encontrado ningún ejemplar ni fotografía que se corresponda con la cita; tampoco en otros herbarios, como B, BM, G, GH, K, LZ y NY, donde hay depositadas colecciones importantes de Bridges. Constance (1965), quien tampoco localizó al ejemplar tipo, hace una detallada reseña histórica y un claro planteo sobre la identidad y el estatus de este taxon, considerando finalmente Azorellopsis trisecta sinónimo de M. pauciflorum —M. albovaginatum var. pauciflorum—. Asimismo, Zech (1992) consideró Azorellopsis trisecta sinónimo de M. albovaginatum var. pauciflorum. Coincidimos con Constance (1965) y Zech (1992) en que Azorellopsis trisecta es una especie de Mulinum. Sin embargo, luego de un cuidadoso y exhaustivo análisis de su descripción original, creemos que este nombre es sinónimo de M. crassifolium y no de M. albovaginatum var. pauciflorum o de cualquier otra especie de Mulinum, sobre la base de las siguientes observaciones: Constance (1965) consideró que el ejemplar de Azorellopsis trisecta citado en el protólogo para Bolivia en realidad fue recolectado en Chile, en Coquimbo o Colchagua; así, infirió que, de las especies que crecen en esta zona, la más afín morfológicamente a lo que describió Wolff era M. pauciflorum y descartó que el ejemplar de Bridges s.n. hubiera sido recolectado en Bolivia. A partir del trabajo de Johnston (1928) sobre las actividades botánicas de Bridges, se conoce que las plantas que recolectó durante el periodo 1844-1847 en Bolivia no fueron enlistadas ni tampoco numeradas. Esto concuerda con lo referido en el protólogo de Azorellopsis trisecta, donde se cita material recolectado por Bridges sin número de recolección. Sin embargo, la única especie de Mulinum que vive en Bolivia es M. triacanthum, morfológicamente muy distinta de lo descrito por Wolff. En definitiva, la etiqueta de Bridges de Bolivia sería dudosa. Sin embargo, después de estudiar el viaje de Bridges por Bolivia, hemos visto que parte de la zona recorrida actualmente pertenece a Chile. En una carta con fecha de mayo de 1844, Bridges dijo haber hecho “(…) una excursión a la parte sur de Bolivia tomando la ruta desde Cobija a Potosí a través del Desierto de Atacama donde también visitó el Valle de Caspana (…)” (Johnston, 1928). Más adelante, en septiembre de 1844, Bridges volvió a Cobija, permaneció allí por unos pocos días y luego se fue a Calama; desde allí, hacia el noreste de Tapaquilcha y luego nuevamente a Potosí (Johnston, 1928). En estas cartas se ve que en 1844 Bridges consideró toda esta región Bolivia. Sin embargo, actualmente la mayor parte del desierto de Atacama, el Valle de Caspana y Calama pertenecen a Chile. Por lo tanto, parte de las recolecciones hechas por Bridges durante el periodo 1844-1845 con etiquetas de Bolivia fueron recolectadas en lo que hoy son las regiones XV de Arica y Parinacota, I de Tarapacá y II de Antofagasta de Chile. En el norte de Chile crecen solo 2 especies de Mulinum, M. crassifolium y M. ulicinum, pero no M. pauciflorum —M. albovaginatum—. Tras realizar el análisis de la descripción original de Azorellopsis trisecta, descartamos que el taxon de Wolff sea sinónimo de M. ulicinum, ya que la planta recolectada por Bridges tiene hojas de 3-4 mm × 1-1,5 mm, crasas, con segmentos foliares de ápice muy obtuso, características que concuerdan perfectamente con la descripción de M. crassifolium y no con la de M. ulicinum. Además es importante destacar que hemos recolectado M. crassifolium durante un viaje por el norte de Chile y el noroeste argentino que incluyó el cruce del desierto de Atacama. Esta especie es abundante y crece en forma de cojines hemisféricos que llaman mucho la atención en esta zona, con lo que consideramos que si Bridges cruzó el desierto de Atacama muy probablemente recolectó M. crassifolium. Teniendo en cuenta todo lo anterior y dado que no se ha podido localizar el material tipo de Azorellopsis trisecta, designamos aquí el pliego recolectado por Calviño 789 (SI) como su neotipo. Este pliego se adecúa a la descripción morfológica del protólogo y fue recolectado al este de Calama, zona que Bridges probablemente visitó.
Además, se designa aquí el pliego recolectado por Philippi s.n., SGO 000003745, como lectotipo de M. crassifolium, porque el otro sintipo corresponde a un pliego destruido de B, del cual solo se dispone de una fotografía (F neg. 003449) y el pliego seleccionado concuerda mejor con el protólogo en lo relativo a la localidad.
Material estudiado.—ARGENTINA. Catamarca: Dpto. Antofagasta de la Sierra, Incahuasi, km 8 camino a Antofagasta, 4200 m, 1-IV-1952, Huek 511 (LIL); Real Grande, 4212 m, 16-I-2003, Pérez 2 (LIL). Salta: Dpto. Los Andes, camino a Mina La Julia, 4500 m, 17-II-1945, Cabrera 8790 (LP); Quebrada del Agua, 3700 m, 9-II-1972, Montaña 532 (LP).
CHILE. Sine loc., desierto de Atacama, 4000 m, II-1988, Philippi s.n. (K 000529786, K 000529787). I Región de Tarapacá: Pcia. Iquique, Collaguasi, Quebrada La Represa, 4550 m, 22-I-1993, Teillier 3028 (CONC). II Región de Antofagasta: Pcia. Antofagasta, Taltal, Cordillera de Volcán Llullaillaco, II-1926, Werdermann 1009 (SI); Quebrada de Las Zorritas, 4450 m, 14-III-1992, Arancio 92-640 (CONC); volcán Llullaillaco, 4500 m, 13-I-1994, Arroyo & al. 94010 (CONC). Pcia El Loa, a 70 km de San Pedro de Atacama por RN 5, a 3 km al sur del desvío a Bolivia, 4460 m, 14-XII- 2010, Calviño & al. 788 y 790 (BCRU); Camino de San Pedro de Atacama al Paso de Jama, 4110 m, 28-XI-2001, Aedo 6997 (CONC); llanos al S del volcán Licancabur, 4050 m, I-1997, Arancio 10647 (CONC); Tolar, Faldas del volcán Licancabur, orillas del Camino Internacional, 4110 m, 24-XI-1996, Rodríguez 3232 (CONC); campamento El Tatio, 4320 m, 5/8-XII-1994, Pedreros 50 (CONC); camino al Tatio, 4500 m, 4-I-1950, Pfister 9374 (CONC); entre el Tatio y Puritama, 4380 m, 23-XI-1996, Rodríguez 3210 (CONC); el Tatio, llaretales cerca de las fumarolas, 4300 m, 16-XII-1964, Schlegel 5124 (CONC); Arroyo Coya, 3780 m, XI-1996, Gutiérrez & López 46 (CONC); Altos de Cablor, 3900-4400 m, 11-XII-1995, Villagrán 8757 (CONC). III Región de Atacama: Pcia. Chañaral, ladera O del Cerro Los Patitos, 4400 m, 24-II-2001, Latorre & al. 206 (CONC); camino entre Salares de Gorbea y La Isla, 4330 m, 22-I-1994, Arroyo & al. 94140 (CONC).
3. M. echegarayi Hieron., Bol. Acad. Nac. Cienc. Córdoba 4: 27 (1881). TIPO: [Argentina:] San Juan, Pie del Tontal y en la Quebrada del Paramillo, I-1876, Echegaray s.n. [lectotipo, designado aquí: CORD 00003539!; isolectotipo: B destruido, foto F neg. 003450!]. Fig. 5.
|
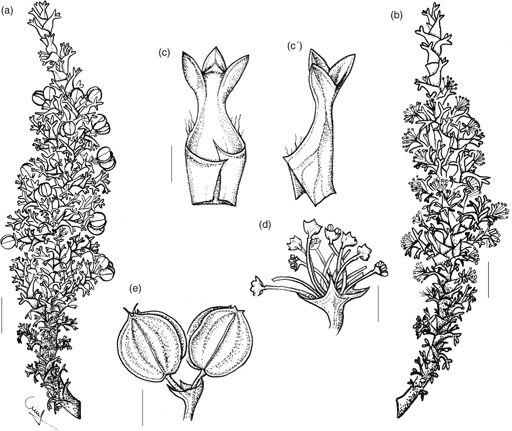 Fig. 5. Mulinum echegarayi: a, rama con flores perfectas; b, rama con flores estaminadas; c, c’, hoja en distintas posiciones; d, umbela con flores estaminadas; e, umbela con frutos [a, e, Semper 11855 (MERL); b-d, Roig & Ruiz Leal 3319 (MERL)]. [Escala: a, b = 1 cm; c = 2mm; d, e = 2,5 mm.] Fig. 5. Mulinum echegarayi: a, rama con flores perfectas; b, rama con flores estaminadas; c, c’, hoja en distintas posiciones; d, umbela con flores estaminadas; e, umbela con frutos [a, e, Semper 11855 (MERL); b-d, Roig & Ruiz Leal 3319 (MERL)]. [Escala: a, b = 1 cm; c = 2mm; d, e = 2,5 mm.]
|
|
Subarbusto androdioico que forma matas de 10-50 cm de altura y 15-20 cm de diámetro, bajas, irregulares, laxas, resinosas, olorosas. Hojas (6)8-10(13) mm, subsésiles, de obovadas a anchamente obovadas, crasas, rígidas; lámina (2)2,5-4 × (3,5)4,5-6(8,5) mm, trisecta o tripartida, con segmento central de (1)2-3(3,5) × (1,2)1,6-2,2(2,6) mm; segmentos de ovados a muy anchamente ovados o triangulares, planos, a veces surcados en la cara adaxial, glabros, con ápice de obtuso a redondeado, mucronado y punzante, con margen entero; pecíolo 1-2,5(3,5) mm, semejante a los segmentos, glabro; base foliar amplexicaule, con margen ciliado. Umbelas que apenas sobrepasan el nivel de las hojas, con 2-5 flores perfectas o 8-17 flores estaminadas; pedúnculos 2-5(6) mm, glabros; involucro de 5 brácteas de 2-3 mm, adpresas, connadas casi completamente en forma de cúpula profunda con lóbulos triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 3-4(4,5) mm, glabros; pétalos 1,5-1,8 × 1,1-1,2 mm, de ovados a anchamente ovados; estilopodio deprimido y estilos 1-3 mm. Fruto (4,5)5-6 × (3)5(6) mm, amarillento, de contorno elíptico a circular, muy comprimido dorsalmente; mericarpio de 1 mm de espesor; alas de (1)1,5-2 mm de anchura, de la misma anchura o más anchas que el cuerpo del fruto, levemente divergentes; carpóforo libre, entero o cortamente bífido.
Distribución geográfica, hábitat y fenología.—Se distribuye por el oeste de Argentina —provincias de San Juan y Mendoza—. Especie endémica de la cordillera del sur de San Juan y el norte de Mendoza, frecuente en la Sierra del Tontal y en cerros relativamente secos cercanos al Cerro Mercedario (Fig. 2). 2000-3800 m s.n.m. Florece y fructifica de enero a principios de abril.
Nombre vernáculo y usos.—Taihuana (Martínez, 2003). Es una planta olorosa y resinosa y se le atribuyen propiedades medicinales para el control de enfermedades hepáticas (Martínez, 2003).
Observaciones.—Esta especie fue dedicada a D.S. Echegaray y su nombre ha sido escrito con una sola i al final del epíteto específico (Hieronymus, 1881; Martínez, 2003) o con dos íes (Martínez, 2008), por lo que es necesario hacer una aclaración. La i griega final de Echegaray es considerada una vocal, por lo que se sigue la recomendación 60C.1.a del CINB (McNeill & al., 2012) para formar la terminación del epíteto basado en un nombre de persona y, por lo tanto, se debe utilizar una sola i final.
El nombre de esta especie fue remitido por Zech (1992) a la sinonimia de M. ulicinum. El aspecto de las ramas de M. echegarayi y M. ulicinum en el material tipo fotografiado es parecido. Sin embargo, aquí se consideran especies distintas. M. ulinum echegarayi se diferencia claramente de M. ulicinum porque este último tiene hojas más grandes —13-17 mm— y segmentos foliares de sección circular y ápice agudo, mientras que M. echegarayi tiene hojas de 8-10 mm y segmentos foliares de sección plana y ápice de obtuso a redondeado.
Designamos aquí el pliego recolectado por Echegaray s.n., CORD 00003539, como lectotipo de M. echegarayi, porque su duplicado era un pliego destruido de B, del cual solo queda una fotografía (F neg. 003450).
Material estudiado.—ARGENTINA. Mendoza: Dpto. Las Heras, Quebrada del Toro, 2800 m, 26-II-1968, Roig s.n. (MERL 45581); Santa Elena, 18 km al N de la villa, adyacencias de la mina de talco “Talcomén”, 2770 m, 1-XII-1984, Ambrosetti 1220 (MERL); 2820 m, 1-XII-1984, Ambrosetti 1243 (MERL); Arroyo Tambillos, 3500 m, 1-IV-1945, Semper 10091 (LIL, MERL); 24 km al NE de Uspallata camino a Villavicencio, 2900 m, 14-I-1963, Boelcke & al. 9940 (SI, BAB); camino de Villavicencio a Uspallata, Antena del Paramillo, 3153 m, 9-XII-2010, Calviño & al. 757 (BCRU); Paramillos de Uspallata, 2700 m, 10-I-1959, Roig & Ruiz Leal 20166 (MERL); Roig 3186 (MERL); 2800 m, 24-II-1983, Roig 11117 (MERL); Guanaco Pampa, 2780 m, 28-I-1960, Ruiz Leal & Roig 20719 (MERL); Roig & Ruiz Leal & al. 3319 (MERL); las minas del Paramillo, 2950 m, 29-I-1897, Kurtz 9463 (BA, CORD). Dpto. Tupungato, Arroyo La Horqueta, 3700-3800 m, 28-II-1949, Semper 11855 (MERL). San Juan: Dpto. Calingasta, Barreal, subida a la antena de El Tontal, 3000 m, 30-XI-2010, Kiesling 10412 (BCRU, MERL); Sierra del Tontal (falda oeste), Estancia El Leoncito, 3000-3300 m, Cocucci 58 (CORD); Cerro Casa de Piedra, 3400 m, 20-II-1984, Kiesling 4532 (SI); Reserva Natural Estricta El Leoncito, 4-IV-2000, Luján 64 (CORD); Quebrada del Telégrafo, 3300 m, 20-III-1999, Luján 46 (CORD); precordillera entre Barreales, Tontal y Retamito, Las Cabeceras, 2020 m, 27-II-1897, Kurtz 9816 (BA, CORD); Cuesta de las Cabeceras, Kurtz s.n. (CORD); subiendo el Cordón del Naranjo por la Quebrada de Las Minas, 18 km al este de la RP 41220-III-1999, Biurrun 5757 (SI); Quebrada del Arroyo del Medio, 11-I-1997, Haene 1625 (SI); La Colorada, Trinchera de San Martín, 7/8-II-1897, Kurtz 9652 (CORD).
4. M. hallei Skottsb., Kongl. Svenska Vetensk. Acad. Handl. 56(5): 278 (1916). TIPO: [Argentina, Santa Cruz:] Puerto Mazaredo in campo, 17-XII-1904, Dusén 5304 (lectotipo, designado aquí: UPS 22 foto!; isolectotipo: K 000529785 foto!). Fig. 6a-d.
|
 Fig. 6. Mulinum hallei: a, fragmento del cojín; b, b’, hoja subsésil y hoja peciolada del mismo individuo; c, flor perfecta; d, umbela con frutos. M. microphyllum: e, fragmento de una mata; f, hoja; g, detalle de la articulación en la base de la lámina; h, flor perfecta; i, umbela con frutos [a-b’, d, Roig 15096 (MERL); c, Losert s.n. (BCRU); e-g, i, Calviño & al. 750 (BCRU); h, Calviño & al. 748 (BCRU)]. [Escala: a, e = 1 cm; b, c, f, h = 2 mm; d, i = 2,5 mm; g = 1 mm.] Fig. 6. Mulinum hallei: a, fragmento del cojín; b, b’, hoja subsésil y hoja peciolada del mismo individuo; c, flor perfecta; d, umbela con frutos. M. microphyllum: e, fragmento de una mata; f, hoja; g, detalle de la articulación en la base de la lámina; h, flor perfecta; i, umbela con frutos [a-b’, d, Roig 15096 (MERL); c, Losert s.n. (BCRU); e-g, i, Calviño & al. 750 (BCRU); h, Calviño & al. 748 (BCRU)]. [Escala: a, e = 1 cm; b, c, f, h = 2 mm; d, i = 2,5 mm; g = 1 mm.]
|
|
Subarbusto andromonoico que forma cojines de 2-8 cm de altura y 10-30 cm de diámetro, chatos, compactos. Hojas (5)7-10(11) mm, subsésiles o pecioladas, de obovadas a angostamente obovadas, coriáceas; lámina 2-3(4) × (2)3-4(6) mm, trisecta —raro tripartida—, con segmento central de 1-2,8(3,5) × (0,9)1-1,3(2) mm; segmentos de ovados a anchamente ovados o angostamente elípticos, planos, surcados en la cara adaxial, glabros, no articulados en su inserción —algunos articulados—, con ápice de obtuso a redondeado, a menudo terminado en un cilio largo blanco, caedizo, no punzante, con margen entero, los laterales raro bipartidos; pecíolo 1-3(4) mm, semejante a los segmentos, glabro; base foliar amplexicaule, con margen ciliado. Umbelas que sobrepasan ampliamente el nivel de las hojas, con 3-20 flores perfectas o 10-20 estaminadas; pedúnculos (6)8-11(16) mm, glabros o, a menudo, con cilios largos y finos esparcidos irregularmente; involucro de 4-6 brácteas de 2,5-4(4,5) mm, reflexas, connadas solo en la base, en forma de cúpula poco profunda con lóbulos angostamente ovados o triangulares, densamente lanosa en el interior, con margen entero o, a menudo, de bi a trilobado, liso o ciliado, con ápice agudo, generalmente terminado en un cilio largo blanco. Flores con pedicelos de (2)2,5-4(6) mm, ciliados en la base; pétalos 1,8-2,1 × 1-1,5 mm, espatulados, marcadamente angostos en la base; estilopodio cónico y estilos 1,6-2 mm. Fruto 5-6 × (4)5-6 mm, de amarillento a rojizo, de contorno anchamente elíptico a circular, muy comprimido dorsalmente; mericarpio de 1-1,5 mm de espesor; alas de (0,5)1 mm de anchura, más angostas que el cuerpo del fruto, paralelas o levemente divergentes; carpóforo libre, entero o cortamente bífido.
Distribución geográfica, hábitat y fenología.—Se distribuye por el sur de la Patagonia argentina —provincias de Chubut y Santa Cruz— (Fig. 2).Habita planicies esteparias, en terrenos arenosos entre piedras sueltas; 50-1200 m s.n.m. Florece y fructifica de noviembre a marzo.
Número cromosómico.—n = 8 (Constance & al., 1971; Constance, 1988).
Observaciones.—La morfología de M. hallei es similar a la de M. microphyllum. Sin embargo, M. microphyllum se diferencia porque forma matas bajas y laxas y tiene hojas de mayor tamaño —11-14 mm—, con segmentos siempre articulados en su inserción, además de pedúnculos mucho más largos —19-30 mm—. Por otro lado, la distribución geográfica de ambas especies solo solapa en la provincia de Chubut. Mulinum microphyllum se distribuye más al norte en Neuquén, Río Negro y Chubut, en zonas altas entre los 800 y 1800 m s.n.m., mientras que M. hallei vive más al sur, entre los 50 y los 1200 m s.n.m. (Fig. 2). Otra característica distintiva es que M. microphyllum tiene un número cromosómico de n = 16 (Constance & al., 1971) y M. hallei de n = 8 (Constance & al., 1971; Constance, 1988).
Skottsberg describió M. hallei sobre la base de la recolección de Dusén 5304 de Puerto Mazaredo —provincia de Santa Cruz— y de su propia colección del Valle del Río Fénix y Lago Buenos Aires —seguramente Skottsberg 855—. En el protólogo de M. hallei, Skottsberg solo incluyó el número de recolección del material de Dusén y no el del suyo. Designamos aquí el pliego recolectado por Dusén 5304, UPS 22, como lectotipo entre los sintipos Dusén 5304 (K 000529785) y Skottsberg 855 (S 10339), porque se encuentra mejor conservado y contiene ramas floríferas y fructíferas que concuerdan con la descripción original y las imágenes de la especie que acompañan el protólogo.
Material estudiado.—ARGENTINA. Sine loc., Patagonia, 1904, Tessleff 5304 (BAB, UC). Chubut: sine loc., 1899, Illín 8 (LP); III-1899, Illín s.n. (BAB 6043, BAB 6045, SI, UC); 17-I-1913, Kraglievich 3949 (BA, UC). Dpto. Escalante, por RN 3 a 600 m desvío a RP 27, camino hacia Río Chico, 639-641 m, 21-I-12, Fernández 20 y 22 (BCRU); Pampa del Castillo, I-1930, Ferruglio 30/1873 (BA, UC); 35 km al N de Comodoro Rivadavia, Cañadón Ferrais, 29-XI-1967, Correa & al. 3985 (UC); Cañadón Ferrais, 17-XII-1929, Ferruglio 30/1878 (BA, UC); 35 km cruce RN 3 camino a Pampa del Castillo, near Comodoro Rivadavia, 03-XII-1967, Correa & al. 3836 (UC); 26-XII-1992, Forcone & al. 10004 y 10005 (UC); 35 km al W del cruce de RN 3 y RP 37, 03-XII-1967, Correa & al. 4063 (BAB, UC); Manantiales Behr, 26-I-1932, Castellanos 6111 (BA, UC); a 29 km de Pampa de Salamanca, 26-XII-1992, Forcone & al. 10000, 10001, 10002 y 10003 (UC); Comodoro Rivadavia, I-1924, sin. col. (LP 22537); Estancia. La Begonia, 30 km al NW de Comodoro Rivadavia, 02-XI-1946, Soriano 2025 (UC); 26-XII-1992, Forcone & al. 10006 (UC); 40 km al NW de Comodoro Rivadavia, Forcone & al. 10007 (UC). Dpto. Languiñeo, Quichaura, Tecka, 11-XII-1946, Soriano 2257 (UC); Tecka, 17-I-1947, Soriano 2426 (UC); 15 km al E de Tecka por RP 6, 1018 m, 19-I-12, Fernández 12 y 13 (BCRU). Dpto. Paso de Indios, RN 24 a 50 km al S de Paso de Indios, desvío a 23 km SO de la ruta, Laguna Blanca, 30-XI-1976, Arroyo & al. 147 (SI); a 51 km SW de RN 24, Estancia La Marfil, 02-XII-1976, Arroyo & al. 210 (SI). Dpto. Rawson, Trelew, Tekachoique, XII-1899, Illín 6038 (BAB, SI); RP 23 S, 28-XI-2002, Bonifacino & Donato 733 (SI). Dpto. Río Senguer, Río Mayo, 14-XII-1981, Cabrera 33177 (SI); 3 km al S de Río Mayo por RN 40, 541 m, 26-I-12, Fernández 34 (BCRU); Valle de la Laguna Blanca, 18-XI-1902, Koslowsky 94 (CORD, SI); Lago Blanco, 23-XII-1902, Koslowsky 12371 (BAB, SI, UC); RP 22 a 2 km al N de Río Mayo, 7-XII-1976, Arroyo & al. 340 (SI); al S de Río Mayo, próximo a RN 40, 560 m, 11-XII-2006, Bonifacino & al. 2971 (SI). Dpto. Sarmiento, 50 km antes de llegar a Sarmiento, 22-I-1962, Ricardi & Matthei 282 (CONC); al NO del Lago Musters, barrancas de cretón en las cabeceras del cañadón Nevazón, 27-XII-1939, Ferruglio 34748 (BA), 28-XII-1939, Ferruglio 34745 y 34747 (BA, UC); 13-I-1940, Ferruglio 34744 (BA, UC); RN 23 a 14 km de Buen Pasto, 03-XII-1976, Arroyo & al. 274 (SI). Dpto. Tehuelches, RN 40, 3 km del centro de Gobernador Costa, 08-XII-2010, Losert 11 (BCRU); 2 km del centro de Gobernador Costa, 09-XII-2010, Losert s.n. (BCRU); 15-XII-2010, Losert s.n. (BCRU); RN 19, a 21 km de NE de Gobernador Costa, 08-XII-1976, Arroyo & al. 421 (SI). Santa Cruz: sine loc., 1899-1900, Ameghino 25 (BA); 7-XII-1945, O´Donell 3936 (LIL); 300 m, 25/26-XI-1928, Donat 60 y 61 (LIL, SI, UC); II-1900, Silvestre 5987 (BAB, SI, UC); III-1898, Venturi 80532 (BAB, SI, UC); I-1914, Hicken & Hauman 360 (SI); Río Santa Cruz (Patagonia austral), II-1950, Silvestre 5988 (BAB, SI, UC); II-1900, Burmeister 6061 (BAB, UC). Dpto. Corpen Aike, Chicorok-Aiken, 29-XII-1898, Burmeister s.n. (BAB 6059, UC); Cte. Piedrabuena, 1-XII-1945, O´Donell 3799 (LIL, UC), 30 km al sur de Piedrabuena por R3, 12-I-1967, Boelcke & al. 12281 (BAB, SI); aprox. 155 km al N de Río Gallegos, ruta 3 a San Julián, 500 m al O de la ruta, 24-I-1989, Zech & Contogiorgakis 21 (CONC, LP, UC); a 160 km de Río Gallegos, 340 m, 13-I-1970, Ruiz Leal & al. 26905 (MERL, UC); Monte León, 27-I-1914, Hicken & Hauman 438 y 445 (SI). Dpto. Deseado, camino Deseado-Cabo Blanco, 19-XI-1963, Correa & al. 2598 (BAB, UC); 3 km de Cabo Blanco a Tellier, 13-XI-1965, Correa & Nicora 3335 (BAB, UC); camino Tellier, paso Gob. Gregores, RP 282, 20-XI-196, Correa & al. 2651 (BAB, UC); ribera N de la ría, Bahía Uruguay, 30 km al W de Puerto Deseado, 18-XI-1963, Correa & al. 2586 (BAB, UC); a 6 k al N de Puerto Deseado, 30-XI-1967, Correa & al. 4011 (UC); Puerto Deseado, Carette s.n. (LP 23378); Née s.n. (MA 476256); 13-I-1914, Hicken & Hauman 138 (SI); 16/17-XI-1945, O’Donell 3560 (LIL, UC); 50 m, 23-I-2012, Fernández 41 (BCRU); RP 18 a 5 km del cruce con RN 26, 262 m, 5-XII-2012, Biganzoli & al. 2380 (SI); Monumento Natural Bosques Petrificados, 5-XI-2002, Raffo 139 (BCRU); 15-XI-1994, Montenegro s.n. (BCRU); 233 m, 27-I-2012, Fernández 40 (BCRU); Cañadón León, XII-1951, Cittadini 52bis (SI); Puerto Deseado, Aeródromo, 10-I-1951, Sleumer 1460 (LIL); estepa del Río Deseado, 25-XI-1979, Roig & Méndez 9887 (MERL); 36 km al O de Las Heras por RN43, 1 km al S de la ruta por camino vecinal o huella, 331 m, 24-I-2012, Fernández 27 (BCRU); Colonia Las Heras, Colonia Pastoril Presidente Carlos Pellegrini, Estancia La Flora, XII-1979, von Thüngen 14 (LP); 13-I-1981, von Thüngen 97 y 103 (LP); Meseta del Lago Buenos Aires, a 10 km de Estancia La Vizcaína a Puesto Lebrum, 1200 m, 23-XII-1968, Ruiz Leal & al. 26425 (MERL, UC); a 15-20 km de Estancia La Aurora, 270 m, 7-XII-1965, Ruiz Leal & al. 24082 (MERL, UC). Dpto. Lago Argentino, Lago Argentino, 1904-1905, Koslowsky 145 (CORD); en las pampas del Río Santa Cruz, 30-X-1874, Berg s.n. (CORD); RN 288, 18 km de Piedra Clavada a Gobernador Gregores, 24-I-1967, Boelcke & al. 12760 (SI). Dpto. Lago Buenos Aires, Skottsberg 855 (S); Perito Moreno, XI-1973, Suárez 821 (SI); a 7,7 km al S de Perito Moreno por RN 40, 595 m, 25-I-2012, Fernández 32 (BCRU); entre arroyo Tecka y Perito Moreno, 640 m, Ruiz Leal & al. 24218 (BAB, UC); RN 40, a 55 km S de Río Mayo, 4-II-1975, Boelcke & al. 16082 (BAB, UC); Cañadón de las Manos Pintadas, al SE de Facundo, 20-I-1972, Llera & Torres 1868-7 y 1921-11 (UC); RP 45 de Perito Moreno a El Portezuelo, km 18, 460 m, 1-XII-2009, Biganzoli & al. 2287 (SI). Dpto. Magallanes, San Julián, 23-XI-1945, O´Donell 3663 (LIL, UC); 14-I-1914, Hicken & Hauman 253 (SI); 50 km north of San Julián, road to Puerto Deseado, 60 m, 30-XII-1938, Eyerdam & al. 23963 (UC); a 70 km antes de Puerto San Julián por RN 3, 11-I-1967, Boelcke & al. 12229 (BAB); Puerto San Julián, XII-1945, Blake 457 (SI); Tres Cerros, Estancia La Lomita, pie del cerro aprox. 3 km, RN 3, 4-XII-1971, Boelcke & al. 15368 (BAB, CONC, UC). Dpto. Río Chico, a 10 km de Las Horquetas hacia Bajo Caracoles, 25-I-1967, Boelcke & al. 12809 (UC); experimental Cañadón León, 23-XI-1955, Soriano 5076 (BAB, UC); 29-XI-1955, Soriano 5098 y 5099 (BAB, UC); entre Estancia Tres Cerritos y Lago Cardiel, 9-XII-1965, Ruiz Leal & al. 24109 (MERL, UC); RN 40, 37 km al N del empalme con la RP 37 a PN Perito Moreno, 661 m, 10-I-2011, Zavala-Gallo & al. 213 (SI).
5. M. leptacanthum Phil., Anales Univ. Chile 85: 709 (1894). TIPO: [Chile, IX Región de la Araucanía:] Nitrito, I-1887, Rahmer s.n. (lectotipo, designado aquí: SGO 000003750 foto!; isolectotipo: SGO 000003749 foto!). Fig. 7.
|
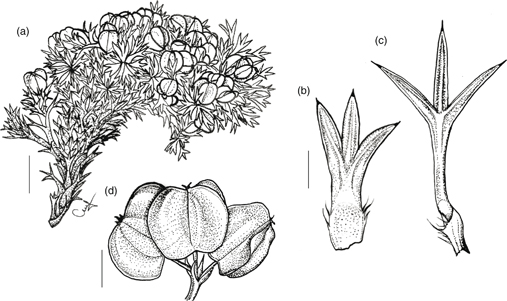 Fig. 7. Mulinum leptacanthum: a, fragmento del cojín; b, c, hojas de distintos cojines; d, umbela con frutos [a, b, d, Ezcurra & al. 2980 (BCRU); c, Marticorena & al. 1398 (CONC)]. [Escala: a = 1 cm; b-d = 2,5 mm.] Fig. 7. Mulinum leptacanthum: a, fragmento del cojín; b, c, hojas de distintos cojines; d, umbela con frutos [a, b, d, Ezcurra & al. 2980 (BCRU); c, Marticorena & al. 1398 (CONC)]. [Escala: a = 1 cm; b-d = 2,5 mm.]
|
|
Azorella nivalis Phil., Anales Univ. Chile 85: 703-704 (1894). TIPO: [Chile, XIV Región de Los Ríos:] Huahuim ad nieves perpetuas, I-1887, Philippi s.n. (lectotipo, designado aquí: SGO 000003662 foto!; isolectotipo: SGO 000003661 foto!).
M. patagonicum Speg., Anales Soc. Ci. Argent. 48: 56-57 (1899). TIPO: [Argentina, Neuquén-Río Negro:] Lago Nahuel-Huapi, Spegazzini s.n. (lectotipo, designado aquí: LP 002799!; isolectotipo: BA 27189!).
Subarbusto androdioico que forma cojines de 3-12 cm de altura y 20-30 cm de diámetro, chatos y compactos. Hojas (9)11-16(20) mm, subsésiles o pecioladas, obovadas, rígidas; lámina (4)5-9(11) × (5)6-11(14) mm, trisecta —raro tripartida—, con segmento central de (3,5)5-7(9) × (1)1,4-2(2,4) mm; segmentos de angostamente ovados a ovados o de angostamente triangulares a triangulares, planos, no surcados en la cara adaxial, glabros, con ápice agudo, espinoso, punzante, con margen entero, los laterales raro bipartidos; pecíolo 2-6(7) mm, poco más ancho que los segmentos en las hojas subsésiles o semejante a los segmentos en las pecioladas, glabro; base foliar amplexicaule —raro poco ensanchada—, con margen ciliado. Umbelas que apenas sobrepasan el nivel de las hojas —raro lo sobrepasan—, con 3-5(7) flores perfectas o (7)9-12(14) flores estaminadas; pedúnculos (2)3-6(10) mm, glabros; involucro de (4)5-6 brácteas de 2-4 mm, adpresas, connadas solo en la base en forma de cúpula poco profunda con lóbulos triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 2-4(6) mm, glabros; pétalos 1,2-2,5 × 1-1,5 mm, de ovados a anchamente ovados; estilopodio de cónico a deprimido y estilos 1-1,5 mm. Fruto 5-7 × 5-7(9) mm, de amarillento a anaranjado-rojizo, de contorno anchamente elíptico a muy anchamente elíptico, poco o muy comprimido dorsalmente; mericarpio de 1-3 mm de espesor; alas de (1)2-2,5(3) mm de anchura, de la misma anchura o más anchas que el cuerpo del fruto, divergentes; carpóforo persistentemente unido a uno de los mericarpios o libre y entero.
Distribución geográfica, hábitat y fenología.—Se distribuye por el noroeste de la Patagonia argentina —provincias de Mendoza, Neuquén y Río Negro— y el sur de Chile —VIII Biobío, IX Araucanía, XI Aysén y XIV Los Ríos— (Fig. 2). Vive en pedreros de alta montaña y cumbres y sitios expuestos; 800-2800 m s.n.m. Florece de finales de diciembre a finales de febrero y fructifica de mediados de enero a finales de abril.
Nombre vernáculo.—Neneo de alta montaña (Argentina; Ferreyra & al., 2006).
Número cromosómico.—n = 8 (Constance & al., 1976; Constance, 1988).
Observaciones.—Se amplía el área de distribución de esta especie en Argentina a la provincia de Mendoza y en Chile a la XI Región de Aysén. Mulinum leptacanthum tiene en esta región chilena una distribución disyunta, no vive en la X Región de Los Lagos, ni en la provincia aledaña argentina de Chubut. Sin embargo, varias de las zonas de altura de la X Región de Chile son de difícil acceso, por lo que no descartamos que también se encuentre en esta región. Por otro lado, el SIB —Sistema de Información de Biodiversidad (Faggi, 1998)— cita la especie para el Parque Nacional Los Alerces, Chubut. Aunque no hay ningún ejemplar de referencia, la presencia de la especie en Chubut también es probable. Por otra parte, en el Catálogo de las Plantas Vasculares del Cono Sur se menciona que M. leptacanthum llega hasta la X Región de Los Lagos de Chile (Martínez, 2008); sin embargo, aquí se ha adoptado la actual división política de Chile, por lo que la región donde crecen estas plantas corresponde a la XIV Región de Los Ríos —antes de 2006 parte de la X Región.
Si bien en un principio se pensó que M. leptacanthum podría ser parte de la amplia variación morfológica observada en M. spinosum, luego de una exhaustiva revisión morfológica del material de ambas especies se pudieron delimitar morfológicamente (Fernández & al., 2017a). Mulinum spinosum tiene mayores umbelas con pedúnculos y pedicelos florales largos —3-6 y 2-4 mm, respectivamente— y hojas también de mayor tamaño —11-16 mm—, con segmentos foliares aciculares, o cuando planos de mayor tamaño —5-7 × 1,4-2 mm— que en M. leptacanthum; los frutos de M. spinosum son menos anchos —5-7 mm— y de contorno de elíptico a circular. Además, M. leptacanthum forma exclusivamente cojines chatos y compactos que no superan los 12 cm de altura, mientras que M. spinosum forma cojines hemisféricos y laxos de más de 20 cm de altura.
Designamos aquí el pliego recolectado por Rahmer s.n., SGO 000003750, como lectotipo de M. leptacanthum, dado que es aquel que contiene una mayor cantidad de material y que concuerda adecuadamente con el protólogo. En cuanto a la designación del lectotipo de Azorella nivalis, se selecciona aquí el pliego recolectado por Philippi s.n., SGO 000003662, por ser aquel cuyo material está mejor preservado y que concuerda bien con el protólogo. Se adoptó este mismo criterio de selección para designar el pliego recolectado por Spegazzini s.n., LP 002799, como lectotipo de M. patagonicum.
Material estudiado.—ARGENTINA. Mendoza: Dpto. Las Heras, Aconcagua, cara E, 3820 m, Miche 171/80/28.3 (UC). Dpto. Malargüe, Las Leñas, 7-XII-1970, Sosa 11 (SI). Neuquén: sine loc., 2-IV-1902, Otto s.n. (BAB 8003, UC). Dpto. Aluminé, Sainuco, 7-XII-1981, Cabrera 32874 (SI); Pampa de Lonco Luan, 1500 m, 4-I-1968, Ruiz Leal & al. 25807 (MERL, UC). Dpto. Chos Malal, extremo NO de la Pampa Ferraína, 30-I-1964, Boelcke & al. 11378 (BAA, BAB, SI, UC); Parque Provincial Tromen, 6-II-1994, Chiapella & al. 223 (BCRU); 2500-2800 m, 29-XII-1999, Ezcurra & al. 2656 (BCRU). Dpto. Huiliches, Parque Nacional Lanín, Volcán Lanín, ladera norte subiendo al refugio por Sendero de Mulas, 1800-1900 m, 1-II-2000, Ezcurra & al. 2980 (BCRU); Bolsico del Diablo, 1500 m, 17-I-1970, Eskuche 928 y 929 (UC); meseta a 1800-1900 m, 1-II-2000, Ezcurra & al. 2974 (BCRU); pedrero a 2100-2350 m, 1-II-2000, Ezcurra & al. 3000 (BCRU); subida al Cerro Los Angeles, 1700-1750 m, 20-II-1974, Correa & al. 5815 y 5817 (BAB, UC); Lago Huechulafquen, subida al Cerro de Los Angeles, 7-II-1948, Donoso & Schwabe 2633 (BAB, UC). Dpto. Lácar, San Martín de los Andes, Cerro Chapelco, 28-I-1973, Cabrera & al. 25964 (SI); 1700 m, 16-II-1968, Cabrera & Crisci 19191 (UC); 1750 m, 26-I-1966, Eskuche 599-19 (UC); del refugio Graef hacia arriba, 3-I-1983, Villamil & al. 2795 (SI); laderas encima del refugio, 1650-1700 m, 23-II-1974, Correa & al. 5853 y 5855 (BAB, UC); filo encima del refugio 1800-1870 m, 23-II-1974, Correa & al. 5912 (BAB, UC); Arroyo Chapelco Grande, arriba de la Laguna La Kika, 1400-1940 m, 31-I-1987, Stuessy & al. 10129 (CONC, UC); 1748 m, 11-III-2008, Calviño & al. 721 (BCRU); cerca del timberline, 30-I-1997, Ferreyra & Chiapella 578 (BCRU); cordón del Cerro Colorado, entre Lago Lácar y Lago Lolog, 1800 m, 25-I-1986, Bernardello & Moscone 629 (CORD); filo del Cerro Colorado, 16-II-19563, Boelcke & Correa 6985, 6987, 6988, 6989 y 6990 (BAB, UC); Hua Hum, Cerro Malo, 2000 m, 19-II-1957, Hunziker 6993 (BAB, UC). Dpto. Loncopué, Chenque Pehuén, cerro al NO del destacamento de gendarmería, 12-I-1984, Rossow & al. 1876 (BAB). Dpto. Los Lagos, Parque Nacional Nahuel Huapi, cumbre del Cerro Pelado, 18-II-1995, Ferreyra 390 (BCRU); filo Cerro Puerto Manzano, 22-II-1953, Boelcke & Correa 7056 (LIL); Puerto Manzano, 1400 m, 24-IV-1968, Diem 3349 (UC); filo del Cerro Colorado, 1800 m, 22-II-1951, Diem 1841, 1844 (LIL); Villa La Angostura, Cerro Bayo, ladera norte, 1430 m, 19-I-1997, Nuñez & Daniels 156 (BCRU); 1600-1800 m, 12-I-2002, Weigend & al. 6864 (BCRU); Río Bonito cerca de Puerto Manzano, 800 m, 23-XI-1947, Diem 1108 (LIL). Dpto. Minas, extremo norte de la laguna Varvarco Campos, cajón Benítez, paso Puerta Vieja, 2600-2750 m, 1-II-1970, Boelcke & al. 14298 (BAA, BAB, SI); paso del Macho, 2280 m, 26-I-1970, Boelcke & al. 13897 (BAA, BAB, SI, UC); sierra de Cochicó, cajón de la Botica, 2500-2700 m, 29-I-1970, Boelcke & al. 14121 (BAA, BAB, SI, UC); cajón del Portillo, paso, 2560 m, 31-I-1970, Boelcke & al. 14179 (BAA, BAB, SI, UC); Reserva Provincial Domuyo, camino geotérmico al sur de Aguas Calientes, Río Covunco, comienzo de la picada sur al Domuyo, 2400-2600 m, 27-I-2001, Ezcurra 3115 (BCRU); valle superior del Arroyo Atreuco, 2010-2050 m, 2-II-1964, Boelcke & al. 11500 (BAA, BAB, SI, UC). Dpto. Ñorquín, Parque Provincial Copahue-Caviahue, pozo geotérmico, ca. Laguna las Mellizas, 2100 m, 30-I-2000, Ezcurra & al. 2909 (BCRU). Copahue, 2050-2150 m, 8-II-1994, Ezcurra 786 (BCRU); 21-XII-1944, Onell 2049 (LIL, SI); serranía rocosa al oeste del pueblo, cerca de curso de agua, 2050-2150 m, 8-II-1994, Ezcurra & al. 799 (BCRU). Dpto. Picunches, RP 13 de Zapala a Aluminé, pasando Primeros Pinos, 1900-2000 m, 1-XII-1999, Ezcurra & al. 2482 (BCRU); Pino Hachado, 11-II-1941, Perez Moreau 45386 (BA, UC); 12-II-1941, Perez Moreau 45379 (BA, UC). Río Negro: Dpto. Bariloche, Bariloche, 18-I-1964, Zöllner 5208 (UC); Parque Nacional Nahuel Huapi, Ferreyra 216 (BCRU); Cerro Challhuaco, Laguna Verde, 26-II-2010, Calviño & al. 746 (BCRU); barranca a orillas de la laguna, 1529 m, 6-III-2008, Calviño & al. 706 (BCRU); sendero a Laguna Verde, 26-II-2010, Calviño & al. 751 (BCRU); Mirador del Ñirihuau, 13-III-2010, Calviño & al. 753 (BCRU); picada a la cumbre, 8-II-1996, Nuñez 47 (BCRU); encima del refugio Neumeyer, 16-XII-1994, Ezcurra & al. 1051 (BCRU); Cerro Catedral, III-1943, Soriano 231 (BAB, UC); 4-III-1943, Perez Moreau 49140 (BA, UC); 6-III-1945, Perez Moreau 49902 (BA, UC); Nido de Cóndores, 6-III-1945, Perez Moreau 49907 (BA, UC); 1700 m, 24-IV-1965, Eskuche 1448 (UC); exposición N, 2000 m, Eskuche 299-1 (UC); 2160 m, 17-I-1946, Montiel s.n. (LIL 246699); 2000 m, 23-I-1995, Ferreyra 342 (BCRU); cordón del Cerro Catedral, Piedra del Cóndor, I-1946, Perez Moreau 50371 (BA, UC); alrededores del refugio Lynch, 2000 m, 4-II-2001, Ezcurra & Quiroga 3163 (BCRU); Cerro Meta, ladera este, 1600 m, 22-XII-1993, Ezcurra & al. 727 (BCRU); 1800 m, 26-I-1995, Ferreyra 330 (BCRU); Cerro Tronador, 27-XII-1974, Rubulis 265a (SI); Cerro Ventana, 23-III-1997, Ferreyra & Siracusa 622 (BCRU); Cerro Cap. De León, Lago Mascardi-NW, 1-II-1970, Eskuche 927b (UC).
CHILE. VIII Región del Biobío: Pcia. Biobío, paso Pichachen, hito fronterizo, 2100 m, 23-I-2001, Baeza & al. 2981 (CONC); camino a laguna Las Totoras, 1580 m, 1-II-2002, Mihoc & al. 4600 (CONC); Sierra de La Polcura, near Laguna de La Laja, 2000 m, 2-II-1968, Zöllner 2293 (UC). IX Región de la Araucanía: Pcia. Malleco, paso Pino Hachado, 1900 m, 2-II-1987, Stuessy & al. 10162 (CONC, UC); 1824 m, 27-XII-1968, Ricardi & Marticorena 5666 (CONC); límite chileno-argentino, 12-I-1994, Villagran & al. 7912 (CONC); 3 km antes de paso Pino Hachado, 1700 m, 10-I-1977, Marticorena & al. 1398 (CONC), 1,7 km al oeste del paso Pino Hachado, 1500 m, 1-III-1962, Greer 1 (CONC); Lonquimay, 2000 m, 9-II-1923, Hollermayer 489 (CONC, UC); 1900 m, 28-II-1936, Hollermayer 1724 (UC); Cerca de Nitrito en la Araucanía, I-1887, Rahmer s.n. (SGO 41607, UC). XI Región de Aysén: Pcia. Aysén, Cerro Pirámide, 1500 m, 19-II-1974, Zöllner 7587 (UC).
6. M. microphyllum (Cav.) Pers., Syn. Pl. 1: 309 (1805). Selinum microphyllum Cav., Icon. 5: 59, lám. 486 fig. 2 (1799), nom. & typ. cons. prop. TIPO: [Argentina, Río Negro, Dpto. Bariloche:] Parque Nacional Nahuel Huapi, Cerro Challhuaco, mirador pedregoso, 1579 m, 26-II-2010, 41°15’29”S; 71°18’05”O, Calviño & Fernández 750 (typ. cons. prop.: SI!; dupl.: BCRU!). Fig. 6e-i.
Huanaca morenonis Kuntze, Revis. Gen. Pl. 3(3): 113 (1898). Diplaspis morenonis (Kuntze) K. Schum., Just’s Bot. Jahresber. 26(1): 367 (1900). M. morenonis (Kuntze) Speg., Anales Mus. Nac. Buenos Aires 7: 295 (1902). Azorella morenonis (Kuntze) Macloskie, Rep. Princeton Univ. Exped. Patagonia, Botany 8: 629 (1905). TIPO: [Argentina, Río Negro o Chubut:] Patagonia, 50/3°, 1882-1884, Moreno & Tonini 368 (holotipo: NY 00405973 foto!).
Subarbusto andromonoico que forma matas de 5-15 cm de altura y 10-30 cm de diámetro, bajas, irregulares, laxas. Hojas (9)11-14(15) mm, pecioladas, obovadas, coriáceas; lámina (3)4(5) × 6-8(9) mm, trisecta, con el segmento central de (3)3,5-4,5(6) × (1,2)1,4-2(2,3) mm; segmentos de angostamente obovados a obovados, planos, surcados en la cara adaxial, glabros, todos articulados en su inserción, articulación a menudo ciliada, con ápice redondeado, terminado en un cilio largo blanco, caedizo, no punzante, con margen entero, los laterales de bipartidos a trífidos —raro enteros—; pecíolo (2)3-6(7) mm, semejante a los segmentos, glabro; base foliar amplexicaule, con margen ciliado. Umbelas que sobrepasan ampliamente el nivel de las hojas, con 2-34 flores perfectas o 10-26 flores estaminadas; pedúnculos (11)19-30(39) mm, glabros o a menudo con cilios largos y finos esparcidos irregularmente; involucro de (4)5-7(8) brácteas de 3-4(5) mm, reflexas, connadas solo en la base en forma de cúpula poco profunda con lóbulos angostamente ovados o triangulares, densamente lanosa en el interior, con margen entero o a menudo bi o trilobado, liso o ciliado, con ápice agudo, generalmente terminado en un cilio largo blanco. Flores con pedicelos de (2,5)3-4,8(5,5) mm, ciliados en la base; pétalos 1,5-3 × 1,2-1,8 mm, espatulados, marcadamente angostos en la base; estilopodio cónico y estilos 1-1,5 mm. Fruto 3-6(7,5) × 3-6(7,5) mm, de amarillento a rojizo, de contorno anchamente elíptico a circular, muy comprimido dorsalmente; mericarpio de 1-2 mm de espesor; alas de 0,5-1 mm de anchura, de la misma anchura o más angostas que el cuerpo del fruto, paralelas; carpóforo libre, entero o cortamente bífido.
Distribución geográfica, hábitat y fenología.—Se distribuye por el noroeste de la Patagonia argentina —provincias de Neuquén, Río Negro y Chubut— (Fig. 2). Valles andinos y planicies altas de mesetas esteparias, generalmente en afloramientos rocosos; 800-1800 m s.n.m. Florece de noviembre a febrero y fructifica de noviembre a fines de marzo.
Nombre vernáculo.—Neneo chico (Ferreyra & al., 2006).
Número cromosómico.—n = 16 (Constance & al., 1971; Constance, 1988).
Observaciones.—Martínez (2008) consideró dudosa la presencia de M. microphyllum en Chile. Lechler recolectó en 1854 tres ejemplares de M. microphyllum que se encuentran depositados en P —P 00115520, P 00115521 y P 03254892—. Las etiquetas de estos ejemplares indican que fueron recolectados en Chile, dos de ellos en pampas de territorio “Pehuelchorum” y el otro en la Cordillera de Ranco. Reiche (1899) en la Flora de Chile también citó material de esta especie de las cordilleras de Valdivia, probablemente a partir de la revisión de los ejemplares de Lechler, ya que el territorio “Pehuelchorum” corresponde al este de la zona valdiviana, tierras donde vivían los pehuelches. Actualmente todas estas localidades de recolección pertenecen a Argentina. Como no se encontró ningún otro espécimen de herbario de esta especie de Chile, se considera que M. microphyllum crece únicamente en Argentina.
Mulinum microphyllum es morfológicamente afín a M. hallei, aunque esta última especie se distingue porque forma cojines chatos, laxos a compactos, las hojas son de menor tamaño —7-10 mm—, con segmentos no articulados en su inserción —a veces articulados en algunas hojas— y pedúnculos mucho más cortos —8-11 mm—. Además, M. hallei vive más al sur, desde Chubut a Santa Cruz (Fig. 2) a menor altitud —50-1200 m s.n.m.— y es diploide, mientras que M. microphyllum vive más al norte, a mayor altitud —desde los 800 a los 1800 m s.n.m.— y es tetraploide.
Como resultado del análisis del protólogo de Selinum microphyllum (M. microphyllum), del pliego original de Cavanilles —Argentina, Santa Cruz, Puerto Deseado, Née s.n. (MA 476256)— y de la ilustración citada en el protólogo, y con el fin de preservar el uso actual del nombre, se ha propuesto la conservación de Selinum microphyllum con un tipo diferente (Fernández & al., 2016a).
En 1898, Kuntze propuso Huanaca morenonis, considerado aquí sinónimo de M. microphyllum, para designar un material recolectado por Moreno & Tonini. Al revisar este material original, observamos que la etiqueta tiene un sello que dice “Patagonia 50/3°, 1882/4 Moreno”, a lo que se agrega, con letra manuscrita, “& Tonini”. Creemos que este sello no se corresponde exactamente con el lugar donde fue recolectado este ejemplar, dado que los viajes de Moreno y Tonini por la Patagonia austral —lat. 50-53º— fueron anteriores a 1880. A partir de este año, Moreno recorrió el lago Nahuel Huapi en Río Negro, y parte de Chubut (Moreno, 1876-1877; Moreno, 1897). Habida cuenta de esto, así como de la falta de recolecciones de la especie al sur del paralelo 46°, consideramos que el holotipo de Huanaca morenonis puede haber sido recolectado en las cercanías de Bariloche o las zonas aledañas de Chubut, no en la Patagonia austral (50/3°).
Material estudiado.—ARGENTINA. Sine loc., territorio Pehuelchorum, XII-1854, Lechler 3000 (P 00115520, P 00115521, UC); Cordillera de Ranco, XII-1856, Lechler s.n. (P 03254892). Chubut: sine loc., Stuckert 18118 (CORD). Dpto. Cushamen, El Maitén, 9-XII-2010, Cottet 8 (BCRU); Epuyén, XII-1925, Pérez Moreau 18663 (BA, UC); Cholila, I-1900, Illín 6058 (SI). Dpto. Escalante, Comodoro Rivadavia, 11-XI-1928, Roivainen s.n. (SGO 79316); 35 km al N de Comodoro Rivadavia, Pampa de Salamanca, directly across highway from Pico Salamanca, 29-XI-1967, Correa & al. 3997 (UC). Dpto. Florentino Ameghino, Uzcudun, 23-XII-1974, Roig & Méndez 15096 (MERL); Garayalde, 26-XII-1992, Forcone & al. 10009 (UC); Estancia La Lochiel, 30 al W de Camarones, 25-X-1946, Soriano 1986 (UC); RN 3, 21 km al S de Garayalde, 28-XI-1967, Correa & al. 3970 (UC). Dpto. Futaleufú, Corcovado, 1901, Illín s.n. (LP 10612); 10/12-III-1901, Illín 257 (SI, UC); Esquel, 10-XI-1949, Soriano 3788 (UC); 15-XII-1970, Garcés 25 (SI); Futalaufquen, 14-XII-1936, Lahitte 52011 (BAB, UC); 24-XII-1941, Künhnemann 552 (BA, UC); a 20 km de Esquel, 16-III-1993, Ayestaran 10008 (UC); Cerro de la Zeta, 4-XII-1941, Künhnemann 471 (BA, UC); Cerro Cuche, Estancia Pampa Chica, 28-I-1947, Soriano 2508 (UC). Dpto. Languiñeo, 10 km al E de Tecka, 13-XII-1981, Cabrera & al. 33116 (SI); 15 km al sur de Tecka, 13-II-1981, Cabrera & al. 33128 (SI). Dpto. Sarmiento, 10 km al W de Hermitte a Sarmiento, 3-XII-1967, Correa & al. 4085 (UC). Dpto. Tehuelches, a 30 km de Gobernador Costa hacia Tecka por RN 40, 6-XII-1967, Correa & al. 4152 (UC). Neuquén: Dpto. Huiliches, Puesto Piedra, camino al Paso Tromen, 17-XI-1969, Eskuche 926 (UC). Dpto Lácar, San Martín de los Andes, 12-XII-1952, Cabrera 11210 (LP, UC); 18/30-I-1959, Ruiz Leal & al. 20244 (MERL, UC); Cerro al N., Dawson 1271 (BAB, CONC, UC). Dpto. Los Lagos, Brazo Huemul, 21-XI-1993, Puntieri 57 (BCRU); sobre costa del Lago Huemul, cerca del Arroyo Quintriqueo, 2-XI-1949, Boelcke & Hunziker 3484 (BAB, UC); Río Limay, alrededores del Anfiteatro, 10-XII-1994, Ezcurra 956 (BCRU); confluencia del Río Limay y el Río Neuquén, XII-1897, Spegazzini s.n. (ex LPS 784 LP); 11 km NE de boca del puente de Río Limay, a 700 m al oeste de RN 40, 10-XII-2010, Tammone 27 (BCRU); entre Rincón de Creide y Cuyín Manzano, 1120 m, 14-XII-1994, Ezcurra & Galende 977 (BCRU); Estancia Fortín Chacabuco, 31-XI-1949, Boelcke & Hunziker 3512 (BAB); XII-1988, Faggi s.n. (BA 78052); alrededor del casco de la Estancia, 31-XI-1949, Boelcke & Hunziker 3572 (BAB, UC); potrero. Zuñiga, 13-I-1958, Boelcke & al. 7574 (BAB). Dpto. Zapala, RP 13 de Zapala a Primeros Pinos, a 10 km de Zapala, 1100-1200 m, 2-XII-1999, Ezcurra & al. 2433 (BCRU); entre Zapala y Laguna Blanca, 915-1300 m, 6-I-1970, Ruiz Leal & al. 26772 (MERL, UC). Río Negro: Dpto. 9 de Julio, Prahuaniyeu, 9-XII-2003, Troiani & Steibel 15715 (SI); Meseta de Somuncurá, inmediaciones de Laguna Paraguay, 1460 m, 20-XII-1967, Ruiz Leal & al. 25591 (MERL, UC); Laguna Azul, Troiani & Steibel 15119 (SI). Dpto. Bariloche, San Carlos de Bariloche, 5-I-1935, Cabrera & Job 39 (LP); 7-I-1947, Teague s.n. (LIL 191353); Los Repollos por el camino a Piedra Pintada, 12-I-1941, Pérez Moreau 45363 (BA, UC); Pampa de Huenuleo, IV-1985, Puntieri 336 (BCRU); base del Cerro León, 8-XII-1986, Puntieri & Chiapella 85 (BCRU); Cerro Runge, XII-1988, Rosso s.n. (BCRU); Los Juncos, 5-II-1942, Pérez Moreau 48689 (BA, UC); Costa Nahuel Huapi, la escuelita, 10-XII-1996, Cordi & al. s.n. (BCRU); Parque Nacional Nahuel Huapi, Cerro Challhuaco, II-1998, Daniels 22 (BCRU); sendero a Laguna Verde, 1579 m, 6-III-2008, Calviño & al. 704 (BCRU); 26-II-2010, Calviño & al. 747 y 748 (BCRU); Valle Challhuaco, Mirador Ñirihuau, 28-I-2001, Stecconi s.n. (BCRU); Valle Challhuaco, Mallín de los Perdidos, ladera NO, 1300 m, 25-I-1997, Nuñez & Amoedo 185 (BCRU); Cerro Otto, 21-I-1945, Soriano 1320 (LP); 1000 m, 7-I-1946, Boelcke & al. 1728 y 1760 (SI, UC); 10-II-1934, Burkart 6642 (BA, UC); 14-I-1942, Pérez Moreau 48694 (BA, UC); 12-II-1943, Pérez Moreau 49142 (BA, UC); on road to Cerro Otto, 1200 m, 10-XII-1971; Arnow 3780 (UC); 1090 m frente al guardaparque del Río Ñirihuau, 10-XII-1996, Ezcurra & Bianchi 1908 (BCRU); Cerro Estratos, ladera este, 1650-1780 m, 25-II-1995, Ferreyra 412 (BCRU); Río Ñirihuau, picada al refugio del Cerro Meta, 1400 m, 22-XI-1993, Ezcurra & al. 703 (BCRU); base del Cerro Ventana, 923 m, 6-XII-2008, Morrone 6135 (SI); Estancia El Cóndor, XII-1988, Faggi s.n. (BA 78048). Dpto. Pilcaniyeu, RN 23 de Bariloche a Pilcaniyeu, Estancia San Ramón, Cerro Villegas, 1021 m, 5-III-2008, Calviño & al. 701 (BCRU); RN 23 a 5 km del cruce de la RN 237, 5 km W of Est. Perito Moreno, 23-XI-1967, Correa & al. 3817 (UC); 10 km al sur de Paso Flores, 10-XII-1981, Cabrera 33021 (SI); Dina Huapi, 30-XI-1987, Puntieri s.n. (BCRU); Comallo, 7-XII-1938, Cabrera 4283 (LP, UC); Colonia Suiza, II-1942, Pérez Moreau 48695 (BA, UC). Dpto. Valcheta, Lago Azul a Cerro Puntudo, 27-XI-2002, Troiani & Steibel 15470 (SI); Cerro Puntudo, 27-XI-2002, Troiani & Steibel 15490 (SI); Cerro Leones, east end of Lago Nahuel Huapi, 800-850 m, 15-XII-1936, West 4757 (UC).
7. M. spinosum (Cav.) Pers., Syn. Pl. 1: 309 (1805). Selinum spinosum Cav., Icon. 5: 59, lám. 487 fig. 1 (1799). TIPO: [Chile, VII Región Maule:] Cordillera del Planchón y Portillo, I/II, Née s.n. (lectotipo, designado aquí: MA 476258 foto!; isolectotipos: CONC 29919, F 68085 foto!, MA 310354 foto!). Fig. 8.
|
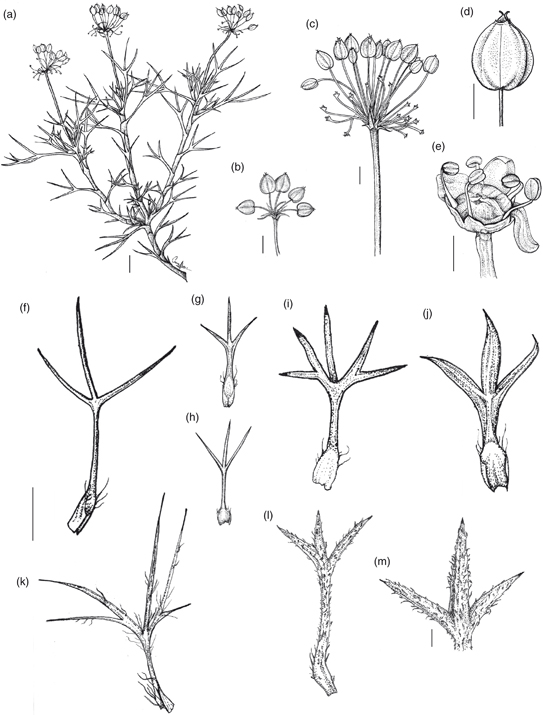 Fig. 8. Mulinum spinosum var. spinosum: a, fragmento de cojín; b, c, umbelas con frutos; d, detalle del fruto; e, flor perfecta en la que se han retirado dos pétalos; f-j, hojas de distintos cojines. M. spinosum var. hirsutum: k, hoja. M. spinosum var. zechii: l, hoja; m, detalle de la pubescencia [a, c, d, f, Calviño & al. 752 (BCRU); b, h, Fernández 7 (BCRU); e, Funes & Arenas 22 (BCRU); g, Fernández 25 (BCRU); i, Arroyo & al. 202079 (CONC); j, Constance & Sparre 3579 (CONC); k, Pérez Moreau 23314 (BA); l-m, Werdermann 268 (SI)]. [Escala: a, f-l = 1 cm; b = 4 mm; c = 5 mm; d, m = 2 mm; e = 1 mm.] Fig. 8. Mulinum spinosum var. spinosum: a, fragmento de cojín; b, c, umbelas con frutos; d, detalle del fruto; e, flor perfecta en la que se han retirado dos pétalos; f-j, hojas de distintos cojines. M. spinosum var. hirsutum: k, hoja. M. spinosum var. zechii: l, hoja; m, detalle de la pubescencia [a, c, d, f, Calviño & al. 752 (BCRU); b, h, Fernández 7 (BCRU); e, Funes & Arenas 22 (BCRU); g, Fernández 25 (BCRU); i, Arroyo & al. 202079 (CONC); j, Constance & Sparre 3579 (CONC); k, Pérez Moreau 23314 (BA); l-m, Werdermann 268 (SI)]. [Escala: a, f-l = 1 cm; b = 4 mm; c = 5 mm; d, m = 2 mm; e = 1 mm.]
|
|
Subarbusto andromonoico o androdioico que forma cojines de 20-100 cm de altura y 20-300 cm de diámetro, hemisféricos, laxos, de un color verde amarillento o a veces glauco grisáceo, muy olorosos. Hojas (9)16-43(51) mm, pecioladas, de angostamente obovadas a obovadas u obtriangulares, rígidas; lámina (7)8-21(24) × 10-28(35) mm, trisecta, con el segmento central de (6)8-19(23) × (0,8)0,9-1,9(2,4) mm; segmentos de angostamente triangulares a subulados, cilíndricos o planos, surcados o no en la cara adaxial, glabros —raro pubescentes—, con ápice agudo, espinoso, punzante, con margen entero —raro los laterales bipartidos—, glabro —raro con pelos—; pecíolo 3-15(20) mm, semejante a los segmentos, glabro —raro pubescente—; base foliar amplexicaule —raro poco ensanchada—, con margen ciliado —raro sin cilios—. Umbelas que apenas o ampliamente sobrepasan el nivel de las hojas, con (2)5-15(18) flores perfectas o 12-28(36) flores estaminadas, a veces con 9-15 flores perfectas centrales y 9-28 flores estaminadas periféricas; pedúnculos 11-43(50) mm, glabros; involucro de (4)5-8 brácteas en 1 ciclo —raro 2—, de (2)2,7-6(7) mm, adpresas o reflexas, connadas solo en la base en forma de cúpula poco profunda, con lóbulos de triangulares a anchamente triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 3-10(13) mm, glabros; pétalos 1,5-2,5 × 1-1,5 mm, ovados; estilopodio de cónico a deprimido y estilos 1-2 mm. Fruto (4)5-7(8) × (3)4-6,5(7) mm, de amarillento a rojizo, de contorno elíptico a circular, poco o muy comprimido dorsalmente; mericarpio de 2-3 mm espesor; alas de 1-2,5(3) mm de anchura, de la misma anchura o más anchas que el cuerpo del fruto, de paralelas a divergentes; carpóforo persistentemente unido a uno de los mericarpios o libre y entero, cortamente bífido o bífido hasta la base.
Distribución geográfica, hábitat y fenología.—Se distribuye por el centro, el oeste, y la Patagonia de Argentina —provincias de San Juan, Mendoza, Neuquén, Río Negro, Chubut, Santa Cruz, San Luis y Buenos Aires— y ampliamente en Chile continental —III Atacama, IV Coquimbo, V Valparaíso, VI Libertador General Bernardo O’Higgins, RM Metropolitana de Santiago, VII Maule, VIII Biobío, IX Araucanía, XI Aysén, XII Magallanes y de la Antártica Chilena— (Fig. 2). Es frecuente en suelos arenosos, roquedales y pedreros, tanto en la estepa como en la alta montaña; 0-4000 m s.n.m. Florece y fructifica de noviembre a marzo.
Nombres vernáculos y usos.—Neneo, chila, uña de gato, coirón, yerba negra y neneo oloroso en Argentina (Cavanilles, 1799; Constance, 1988; Martínez, 2003; Estomba & al., 2005; Ferreyra & al., 2006); hierba negra, dichillo, hierba de la culebra, matabarrosa y neneo en Chile (Muñoz & al., 1999).
Se le atribuyen propiedades medicinales analgésicas, urinarias y digestivas (Niemeyer, 1995; Muñoz & al., 1999; Estomba & al., 2005; Barboza & al., 2009). Forma parte importante de la dieta de ovejas y cabras en la región patagónica y el exudado resinoso que libera produce un sabor amargo en la carne de los animales que la consumen (Cavagnaro & al., 2003; Seoane & al., 2011).
Número cromosómico.—n = 8, 16, 24? (Constance & al., 1971; Constance, 1988).
Observaciones.—Este taxon tiene la distribución más amplia, tanto en longitud como en latitud geográfica y uno de los más recolectados del género. Mulinum spinosum es muy conocido como componente principal de la estepa herbáceo-arbustiva de la Patagonia y el Monte austral. Muestra una gran variación ecológica y morfológica y, como consecuencia, se ha generado una gran cantidad de nombres para el mismo.
Zech (1992) en su tratamiento del género consideró M. spinosum una especie con 4 variedades que se diferencian por la pubescencia, el tamaño de las hojas y si los segmentos son más o menos aciculares. Posteriormente, Zech (1995) validó solo una de estas variedades, M. spinosum var. minus (Kuntze) Zech, que incluye individuos de hojas pequeñas y glabras. Sin embargo, la observación en el campo de esta especie a lo largo de casi toda su distribución, el análisis de abundante material de herbario y los estudios morfométricos, han permitido observar una gradación continua del tamaño de las hojas y las umbelas que está principalmente asociada a la longitud geográfica (Fernández & al., 2017a). Consideramos, pues, que no se justifica la división de esta especie en variedades de acuerdo con estos caracteres. Por el contrario, se han observado poblaciones chilenas de Huasco —III Región de Atacama— y en algunas otras de Colchagua —VI Región del Libertador General Bernardo O’Higgins— una variación del tipo de indumento, carácter que falta en individuos de los demás sitios del área de distribución de la especie. A su vez, el tipo de indumento de las plantas es distinto entre las regiones III de Atacama y VI del Libertador General Bernardo O’Higgins. Así, se han observado dos caracteres cualitativos asociados a áreas geográficas particulares y, por lo tanto, reconocemos sendas variedades de M. spinosum.
Designamos aquí al pliego MA 476258 como lectotipo de Selinum spinosum entre todos los sintipos por ser el que presenta mayor abundancia de material y porque se ajusta bien al protólogo —hojas cilíndricas y planas, flores estaminadas y perfectas, y frutos circulares.
Clave para determinar las variedades de M. spinosumTOP
| 1. |
Hojas glabras, con indumento solo en la base foliar |
7a. var. spinosum |
| – |
Hojas pubescentes |
2 |
| 2. |
Hojas con pocos pelos muy largos y finos, distribuidos irregularmente por la hoja; Chile, región VI, provincia de Colchagua |
7b. var. hirsutum |
| – |
Hojas con abundantes pelos largos y finos distribuidos por toda la hoja y que confieren una coloración grisácea; Chile, región III, provincia de Huasco
|
7c. var. zechii |
7a. M. spinosum var. spinosum. Fig. 8a-j.
Selinum proliferum Cav., Icon. 5: 59, lám. 486 fig. 1 (1799). M. proliferum (Cav.) Pers., Syn. Pl. 1: 309 (1805). Bolax prolifer (Cav.) Spreng., Syst. Veg. 6: 361 (1820). M. spinosum var. proliferum (Cav.) Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Argentina, Santa Cruz:] Puerto Deseado, XII, Née s.n. (lectotipo, designado aquí: MA 476257 foto!).
M. echinus DC., Prodr. 4: 79 (1830), syn. nov. TIPO: Chile, Haenke s.n. (lectotipo, designado aquí: PR 612387 foto!; isolectotipo: PRC 452781 foto!).
M. laxum Phil., Linnaea 33: 90 (1864). TIPO: [Chile, VIII Región Biobío:] In Andibus Antuco, I-1839, Gay 1513 (holotipo: SGO 000003748 foto!).
M. leoninum Lorentz in Roca, Exped. Río Negro, Bot.: 222, lám. 6(1) figs. 1-5 (1883). M. spinosum var. leoninum (Lorentz) Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Argentina, Buenos Aires:] cerca de Leones, entre el Fuerte Argentino y Nueva Roma, 4-V, Lorentz s.n. [lectotipo, designado aquí: Roca, Exped. Río Negro, Bot., lám. 6(1) fig. 1 (1883)].
M. chillanense Phil., Anales Univ. Chile 85: 707 (1894). TIPO: [Chile, VIII Región Biobío:] Valle de las nieblas, I-1877, Philippi s.n. (lectotipo, designado aquí: SGO 000003741 foto!; isolectotipo: SGO 000003742 foto!).
M. ovalleanum Phil., Anales Univ. Chile 85: 709 (1894). TIPO: [Chile, IV Región Coquimbo:] Cerro Huatulame, I-1890, Geisse s.n. (lectotipo, designado aquí: SGO 000003752 foto!; isolectotipo: SGO 000003751 foto!).
M. spinosum (var. leoninum) f. toscae Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Argentina, Buenos Aires:] Toscahügel Leones am Río Sauce Chico, 4-V-1879, Lorentz s.n. (lectotipo, designado aquí: CORD 2485!).
M. spinosum var. quinquepartitum Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Chile, V Región Valparaíso:] Paso Cruz 34°, Cordillera, 1500-2000 m, Kuntze s.n. (holotipo: NY 00038521 foto!).
M. spinosum (var. triacanthum) f. minus Kuntze, Revis. Gen. Pl. 3(3): 114 (1898), syn. nov. M. spinosum var. minus (Kuntze) Zech, Kurtziana 24: 192 (1995). TIPO: [Argentina, Río Negro o Chubut:] Patagonia 50/3°, 1882/4, Moreno & Tonini 372 (holotipo: NY 00073905 foto!).
M. spinosum var. trispinescens Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Argentina, Mendoza:] 34º Paso Cruz, 2000 m, Kuntze s.n. (lectotipo, designado aquí: NY 00073906 foto!).
M. patagonicum Gand., Bull. Soc. Bot. France 59: 710 (1913). M. gandogeri M. Hiroe, Umbell. World: 1732 (1979). TIPO: [Argentina, Santa Cruz:] Patagonia, Lago San Martín, 9-I-1909, Skottsberg 706 (lectotipo, designado aquí: LY, foto!; isolectotipo: SGO 000003754 foto!).
M. spinosum var. longeinvolucratum Hicken, Darwiniana 1(2): 61 (1923). TIPO: [Argentina, Mendoza:] Arroyo Blanco, Río Atuel, XII-1912, Gerth 58bis (holotipo: SI 00738!).
Hojas glabras, con segmentos foliares surcados o no en la cara adaxial y margen entero.
Distribución.—Se distribuye por Argentina —provincias de San Juan, Mendoza, Neuquén, Río Negro, Chubut, Santa Cruz, San Luis y Buenos Aires— y Chile —IV Coquimbo, V Valparaíso, VI del Libertador General Bernardo O’Higgins, RM Metropolitana de Santiago, VII Maule, VIII Biobío, IX Araucanía, XI Aysén, XII Magallanes y de la Antártica Chilena— (Fig. 2). 0-3300 m s.n.m.
Observaciones.—Se amplía el área de distribución de M. spinosum var. spinosum a la provincia de San Luis en Argentina. A pesar de haber encontrado un único ejemplar recolectado en esta provincia —en la Sierra del Morro—, el material concuerda perfectamente con las características morfológicas descritas para la variedad.
El concepto de M. echinus de Constance (1988) se basa en un ejemplar recolectado por Haenke en Chile y erróneamente etiquetado como isotipo de M. echinus por Mathias y Constance en 1969-1970. Con este concepto, M. echinus correspondería a plantas más pequeñas con hojas e inflorescencias también menores que las de M. spinosum. Estas plantas también se diferencian en el campo por su color glauco grisáceo y el fuerte aroma que despiden las hojas al romperse (Constance, 1988; Martínez, 2003). Sin embargo, al analizar el protólogo de M. echinus descubrimos que el ejemplar de Haenke, hasta ahora considerado isotipo de M. echinus, no pertenece al material tipo de la especie, dado que los datos de la etiqueta no coinciden completamente con los datos del protólogo. En cambio, se encontró el material tipo de M. echinus: PR 612387 y PRC 452781. Morfológicamente estos ejemplares son similares a los ejemplares típicos de M. spinosum. Por lo que, sobre la base de lo expuesto arriba y tras los pertinentes análisis morfométricos y filogenéticos (Fernández & al., 2017a, 2017b), M. echinus pasa a ser sinónimo de M. spinosum var. spinosum. Estos resultados además señalan que tampoco merece la pena utilizar un nuevo nombre para los individuos morfológicamente similares a lo que fue erróneamente considerado isotipo de M. echinus, ya que también entran dentro del rango de variación de M. spinosum.
En cuanto a la entidad M. spinosum f. minus, Martínez (2008) lo consideró sinónimo de M. triacanthum. Sin embargo, el ejemplar tipo de esta forma fue recolectado en la Patagonia austral —Río Negro o Chubut, Argentina—, área a la que M. triacanthum no llega y, además, tiene hojas pequeñas como las que se observan en individuos de M. spinosum de Chubut o de Santa Cruz y nunca en M. triacanthum.
Designamos aquí el pliego recolectado por Née s.n., MA 476257, como lectotipo de Selinum proliferum porque es el único que coincide con el protólogo. La ilustración que acompaña el protólogo corresponde a un ejemplar anómalo. Se designa el pliego recolectado por Haenke s.n., PR 612387, como lectotipo de M. echinus, ya que su duplicado presenta menos material. Para M. leoninum se designa como lectotipo la ilustración de un ejemplar —Roca, Exped. Río Negro, Bot., lám. 6(1) fig. 1 (1883), icon— que acompaña al protólogo, ya que no se ha localizado ningún otro material tipo. Dicha figura, además de coincidir perfectamente con el protólogo, ilustra abundante material vegetativo y una umbela florífera pedunculada. También se designan dos pliegos recolectados por Philippi s.n., SGO 000003741 y SGO 000003742, en el Valle de las Nieblas, como lectotipo e isolectotipo de M. chillanense, respectivamente, ya que están mejor conservados que el material sintipo de los Baños de Chillán, SGO 41613. Para M. ovalleanum se designa el pliego recolectado por Geisse s.n., SGO 000003752, como lectotipo entre su duplicado SGO 000003751 y el pliego SGO 000003753, por ser un ejemplar completo con umbelas floríferas y fructíferas —en SGO 000003751 hay solo umbelas floríferas y en SGO 000003753 también hay umbelas fructíferas pero no conectadas, por lo que posiblemente provengan de otro ejemplar—. Mulinum spinosum (var. leoninum) f. toscae se lectotipificó con el pliego recolectado por Lorentz s.n., CORD 2485, ya que presenta abundante material vegetativo y una umbela florífera cortamente pedunculada, que concuerda con la diagnosis de la forma. Además, la etiqueta original de Lorentz dice “Mulinum toscae n. sp.” que concuerda con la observación del protólogo que indica ”Lorentz ined.”. Mulinum spinosum var. trispinescens se lectotipificó con el pliego recolectado por Kuntze s.n., NY 00073906, ya que presenta más material que el de Moreno & Tonini 371, NY 00038520. Para M. patagonicum se designa al pliego de LY como lectotipo ya que presenta buen material para su reconocimiento y etiquetas originales que se ajustan mejor al protólogo que el pliego de SGO.
Material estudiado.—ARGENTINA. Sine loc., Patagonia, Moreno & Tonini 371 (NY 00038520). Buenos Aires: Partido Bahía Blanca, Tornquist a Bahía Blanca, RN 33, km 38, 9-XII-1970, Burkart & Múlgura 28191 (SI, UC); Partido Saavedra, Sierra de Cura Malal, 4-XI-1939, Cabrera 5507 (SI). Partido Tornquist, Choique, 7-XII-1940, Cabrera 6720 (SI); Sierra de la Ventana, 11-XI-1907, Hicken 116323 (SI); Sierra de la Ventana, Cerro de la Ventana, 17-II-1940, Cabrera 5767 (SI); Paraje ”La Gruta”, 450 m, IX-1984, Bottino 247 (MERL). Partido Patagones, a 25 km al SE de Carmen de Patagones, 26-XI-1963, Fabris & Schwabe 4918 (LP). Partido Saavedra, Sierra de Curamalal, 4-XI-1979, Cabrera 5507 (LP). Chubut: Dpto. Biedma, Península Valdés, 17-II-1938, Scott de Birabén 450 (LP); Istmo Carlos Ameghino, 76 m, 22-XII-1967, Ruiz Leal & al. 25663 (MERL, UC); 40 km de Puerto Madryn a Puerto Pirámides (Península Valdés), 26-XI-1967, Correa & al. 3936 (UC). Dpto. Cushamen, 11 km después de Leleque, 12 km al N, toward El Maitén, 8-XII-1967, Correa & al. 4181 (UC). Dpto. Escalante, Pampa del Castillo, 24-II-1938, Scott de Birabén 518 (LP); entrada a Pico Salamanca, 389 m, 19-XII-2011, Urtubey & Baztarrica 634 (SI); 35 km cruce RN 3 camino Pampa del Castillo, 3-XII-1967, Correa & al. 4065 (UC). Dpto. Florentino Ameghino, aprox. 8 km al S del ingreso a Camarones por RN 3, 381 m, 21-I-2012, Fernández 17 (BCRU); 35 km al sur de Garayalde, 28-XI-1967, Correa & al. 3972 (UC); RN 3, aprox. a 80 km al norte de RP 30, 24-I-1984, Rossow & al. 2374 (BAB). Dpto. Futaleufú, zona basal de la vertiente norte de Nahuel Pan, al E de Esquel, 6-I-1974, Sermolli & Bizzarri 7372 (CONC). Dpto. Languiñeo, 15 km al E de Tecka por RP 62, 1018 m, 19-I-2012, Fernández 14 (BCRU). Dpto. Mártires, Gaimán, Las Chapas, 27-XI-1976, Arroyo & al. 20 (SI). Dpto. Paso de los Indios, Valle de las Ruinas, 19-XII-1981, Cabrera 33279 (SI); RN 24, entre Paso de los Indios y Sarmiento, 838 m, 17-XII-2011, Urtubey & Baztarrica 579 (SI). Dpto. Rawson, 10 km al sur de Trelew, 18-XII-1981, Cabrera 33270 (SI). Dpto. Río Senguer, praderas altas del Cañadón grande, Vuelta del Senguer, 10-II-1927, Cabrera 34 y 35 (LP); Valle de la Laguna Blanca, 26-XII-1901, Koslowsky 95 (CORD, SI). Dpto. Sarmiento, 25 km del cruce de Sarmiento hacia Pampa de María Santísima, Lago Musters, 4-XII-1967, Correa & al. 4096 (BAB, UC); 47 km al norte de Sarmiento, 3 km NW of Los Manantiales, Sarmiento-Facundo, 6-XII-1967, Correa & al. 4136 (UC). Dpto. Telsen, a 35 km NO de Telsen, camino a Cerro Cayuqueo, 11-XII-1976, Arroyo & al. 462 (SI). Mendoza: sine loc., Stuckert 2834 (CORD); 15-I-1886, Kurtz 3387 (CORD, UC). Dpto. Las Heras, Pta. de las Vacas, 4-II-1939, Troncoso 9310 (SI); III-1901, Spegazzini s.n. (SI 6911); cercano a Uspallata, Los Hornillos, 5-II-1987, Naranjo & al. 906 (SI); Quebrada del Toro, Cerro Pelado, 2400 m, 18/22-I-1897, Kurtz 9372 (CORD); Pto. Chacay, 2475 m, 18-I-1985, Martínez Carretero 144 (MERL); Sierra de Uspallata, paramillos al N de la Quebrada, 2600 m, 18-XII-1984, Roig & al. 46839 (MERL). Dpto. Luján de Cuyo, Potrerillos, El Salto, estribaciones del Cerro Cabras, 1780-1830 m, 11-I-1984, Del Vitto & al. 36530 (MERL); Vallecitos, cerca del refugio Edelweiss, 2826 m, 11-II-2003, Cocucci & al 2211 (CORD); Agua de La Pampa, Pampa de la Polcura, 2550 m, 28-I-1962, Ruiz Leal & al. 21988 (MERL, UC); Cerro de la Polcura, 28-I-1962, Roig 4232 (MERL); Valle del Río Tupungato, Río Tupungato, 2452 m, 27-XII-2006, Méndez 9888 (MERL). Dpto. Malargüe, Agua del Choique, 2170 m, Ruiz Leal & al. 22387 (MERL, UC); camino a límite paso Pehuenche, base de ladera S, 1825 m, 8-III-2007, Méndez 10462, 10473 (CORD, MERL); 5 km al NE de Valle Hermoso camino a Los Molles, 28-I-1963, Boelcke & al. 10309 (BAB, SI, UC); 1700 m, 17-I-1982, Cabrera 33331 (SI); 5 km de Los Molles hacia Las Leñas, 14-I-1989, Zech & Contogiogakis 2 (LP, UC); Valle del Río Tordillo, 2220 m, 15-I-1989, Zech & Contogiogakis 6 y 7 (CONC, LP, UC); 15 km de Valle Hermoso hacia Las Leñas, 15-I-1989, Zech & Contogiogakis 8 (LP, UC); Valle del Salado, Alto de los Caracoles, “Camino del Sol”, 1750 m, 23-XII-1984, sin colector (SI 165407); Valle de Las Leñas, 2250 m, 8-XII-1985, sin colector (SI 165410); 9-XII-1985, sin colector (SI 165409); entre Barrancas y Ranquil Norte, 1276 m, 2-XII-2007, Cocucci 4132 (CORD); camino a Reserva La Payunia, 6 km de la pasarela, 1257 m, 2-XII-2007, Cocucci 4133 (CORD); RN 40, 35 km al sur de Malargüe, 1917 m, 2-XII-2007, Cocucci 4134 (CORD); Valle del Atuel, 1840-1920 m, Ruiz Leal & al. 23156 (MERL, UC). Dpto. San Carlos, RN 40, 126 km al sur de Tunuyán, Arroyo Hondo, 23-I-1963, Boelcke & al. 10205 (SI, UC); 13,5 km de RN 98, hacia Lago Diamante por RN 101, 2040 m, 30-I-2002, Chandler & Bayer 1111 (LP). Dpto. San Rafael, Valle del Río Atuel, 78 km al oeste de El Sosneado, camino planta mina de azufre, 26-I-1963, Boelcke & al. 10236 (BAB, SI, UC); Laguna El Sosneado, 25-I-1964, Roig 4855 (MERL); Laguna La Niña Encantada, 19-II-1941, Dawson 1032 (LP); Valle de Las Leñas, Ao. De Las Leñas, 2030 m, 30-I-1966, Ruiz Leal & al. 24544 (MERL); Sierra del Nevado, Arroyo Chacay-Có, puesto Barroso, 1300 m, 3-XII-1973, Boelcke & al. 15538 (BAA, BAB, MERL, SI, UC); Sierra del Nevado, pampa entre Arroyo Agua del Guacho y el zanjón del plateado al S-SE de los Cerros Morados, 8-XII-1973, Boelcke & al. 15747 (MERL, UC); Cuadro Benegas, W del Cerro La Leña, 2325 m, Méndez & Wuilloud 32975 (MERL); Cerro Nevado, 23/25-XII-1887, Kurtz 5622 (CORD); norte de Cerro El Rodeo, oeste 25 de Mayo, 1700 m, 1976, sin colector (SI 165406); Portezuelo del Calqueque, 28-I-1888, Kurtz 5904 (CORD); Río Salado superior, Cerro de Los Molles, 7-I-1893, Kurtz 7505, 7507 (CORD); Río Salado superior entre Arroyo Vertientes Amarillas y Arroyo Los Moros, 1940 m, Ruiz Leal & al. 24532 (MERL, UC). Dpto. Tunuyán, Los Árboles, 1240 m, 25-I-1934, Ruiz Leal & al. 2021 (MERL). Dpto. Tupungato, camino a La Carrera por Potrerillos, 2300 m, Ruiz Leal & al. 23719 (MERL, UC); La Media Luna (Estancia Lo Vila), 7-I-1935, Ruiz Leal & al. 2887 (MERL). Neuquén: sine loc., 1-I-1900, Hicken 6081 (SI). Dpto. Aluminé, Aluminé, Cerro Batea Mahuida, 1753 m, 13-III-2008, Calviño & al. 726 y 727 (BCRU); RP 23, entre Paso Pino Hachado y Villa Pehuenia, 1384 m, 19-I-2003, Cocucci & Sérsic 2362 (CORD). Dpto. Catán Lil, Sierra de Catán Lil, bajada a 1° de Mayo, 9-XII-1981, Cabrera 32965 (SI). Dpto. Confluencia, Plaza Huincul, 3-XII-1943, Plotrick 49 (LP). Dpto. Chos Malal, RN 40, entre km 2243 y 2242, 1025 m, 1-XII-2007, Cocucci 4124 (CORD); 40 km al S de Chos Malal, 18-XI-1967, Cabrera 18713 (LP, UC); a 9 km de Chos Malal camino a Andacollo, 819 m, 12-I-1964, Boelcke & al. 10648 (BAB, SI, UC); RN 40, 17 km al norte de Chos Malal, camino a Buta Ranquil, 1300 m, 5-II-1994, Chiapella & al. 122 (BCRU); 36 km al norte de Chos Malal, cerca de la Laguna Aquincó, 5-II-1994, Chiapella & al. 144 (BCRU); NE de Chos Malal, 1750 m, 16-I-1995, Ezcurra & al. 1259 (BCRU); Parque Provincial Tromen, Volcán Tromen, quebrada del arroyo que va a la laguna del volcán, 2360 m, 6-II-1994, Chiapella & al. 192 (BCRU); entrada al parque, ladera sur, 1910 m, 15-I-1995, Chiapella & al. 444 (BCRU); planicie frente a la laguna del volcán Tromen, 2135 m, 16-I-1995, Chiapella & al. 528 (BCRU). Dpto. Collón Curá, Collón Curá, 6-XII-1972, Latour 205b (BCRU); RN 237, embalse Alicurá, 726 m, 30-XI-2007, Cocucci & Sérsic 4120 bis (CORD); RN 40, unos 5 km al norte del puente Collón Curá, 604 m, 30-XI-2007, Cocucci & Sérsic 4121 (CORD); 20 km de Piedra del Águila en dirección a Bariloche, 13-XII-2009, Bach 304 (BAB). Dpto. Huiliches, 13 km al sur de Junín de los Andes, near Río Quilquihue, 12-XII-1967, Correa & al. 4202 (UC). Dpto. Lácar, Alicurá, XII-1979, González 210 (LP); alrededores del lago Currué Chico, 910 m, 23-II-2005, Barboza & al. 1207 (CORD). Dpto. Los Lagos, Estancia Fortín Chacabuco, 14-XI-1950, Boelcke & al. 4301 (BAB); Parque Nacional Nahuel Huapi, RP 65, camino a Villa Traful, a 600 m del desvío Confluencia-RN 237, 28-XI-2006, Morrone 5702 (SI); 400 m al NO de RN 40, 11 km NE del puente boca del Río Limay, 780 m, 26-XII-2010, Fernández 6 (BCRU); entre Rincón Grande y Rincón Chico, RN 237, sobre la ruta junto al Río Limay, 710 m, 20-XII-2002, Cocucci & al. 2112 (CORD); Confluencia, 10-XII-1944, Descole 2420 (LIL); Parque Nacional Nahuel Huapi, margen del río en la Segunda Angostura del Limay, 6-X-2003, Ezcurra 3502 (BCRU); 2,3 km al O de RN 40, 1180 m, 27-XII- 2010, Fernández 7 (BCRU); entre Rincón de Creide y Cuyín Manzano, 1120 m, 14-XII-1994, Ezcurra & Galende 1008 (BCRU). Dpto. Minas, Andacollo, Mina Guaraco, 8-XII-1952, Cabrera 11132 (LP, UC); Pichi-Neuquén, puesto de Gendarmería, 21-I-1970, Boelcke & al. 13564 (BAA, BAB, SI, UC); confluencia de los ríos Pichi-Neuquén y Neuquén, Cerro de las Yeguas, 23-I-1970, Boelcke & al. 13761 (BAA, BAB, SI); laguna Varvarco, extremo sur, 28-II-1970, Boelcke & al. 13972, 13974 (BAA, BAB, SI, UC); extremo norte de la laguna Varvarco Campos, cajón Benítez, curso medio, 31-II-1970, Boelcke & al. 14246 (BAA, BAB, SI); Lagunas de Epulafquen, cerro al NO de las lagunas, 17-I-1964, Boelcke & al. 10970 (BAA, BAB, SI, UC). Dpto. Ñorquin, RN 40, km 2576-2575, al sur de Chos Malal cerca de Nanauco, 955 m, 1-XII-2007, Cocucci 4127, 4128 (CORD); Parque Provincial Copahue-Caviahue, 22-II-2003, Canessa 53 (BCRU); termas de Copahue, 17-II-1940, Cabrera 6177 (LP); Salto del Agrio, camino al mirador del cañadón, 1490 m, 15-III-2008, Calviño & al. 734 (BCRU). Dpto. Pehuenches, RN 40 de Chos Malal, 1203 m, 2-12-2007, Cocucci 4131 (CORD); entre Balsa Huitrin y Buta Ranquil, 16-XII-1965, Ruiz Leal & al. 24357 (MERL, UC). Dpto. Zapala, Zapala, 1000 m, 5-XII-1952, Cabrera 11067 (LP, UC); cerca de Zapala, 6-XII-1981, Cabrera 32836 (SI); RN 40, unos 20 km al sur de Zapala entre km 2243 y 2242, 1090 m, 1-XII-2007, Cocucci 4125 (CORD); Pino Hachado, 14-II-1968, Cabrera & Crisci 19112 (LP, UC); Parque Nacional Laguna Blanca, 200 m al E de seccional La Vega, 1,2 km al E, 2,1 km al N del Cerro de la Laguna, 24-I-2006, Sage 1765128 (BCRU); lado este de Laguna Verde, 14-I-2005, Sage 1703210 (BCRU); Cerro Lotena, 850 m, 8-I-1966, Ruiz Leal & al. 24437 (MERL, UC); alrededores del Cerro Lotena, laderas exposición E, 20-XI-1983, Boelcke & al. 8818 (BAB). Río Negro: Dpto. Bariloche, Bariloche, Colonia Suiza, 15-XI-1996, Cordi & al. s.n. (BCRU); entre Arroyo Casa de Piedra y Lago Moreno, 10-III-2012, Fernández 44 (BCRU); Parque Nacional Nahuel Huapi, Ferreyra 216 (BCRU); Río Ñireco, 24-I-2002, Ezcurra & al. 3291 (BCRU); Cerro Challhuaco, 6-III-2008, 1542 m, Calviño & al. 709 (BCRU); Laguna Verde, 26-II-2010, Calviño & al. 745 (BCRU); picada a la cumbre, pedrero ladera NE, 1600-1800 m, 16-II-1996, Nuñez & Ezcurra 84 (BCRU); Cerro Catedral, 11-III-1973, Gómez 205a (BCRU); 26-II-2010, Calviño & al. 752 (BCRU); 2000 m, 25-II-1997, Ferreyra & Ezcurra 610 (BCRU); Cerro Meta, ladera oeste, 1700-1750 m, 12-I-1994, Ferreyra & Clayton 198 (BCRU); ladera norte y oeste, 1700 m, 13-II-1994, Ferreyra & Clayton 291 (BCRU); valle del Río Manso Inferior, 9-II-1998, Funes & Arenas 22 (BCRU); Cerro López, 1542 m, 12-II-1994, Chiapella & Clayton 625 (BCRU); Nacimiento del Río Limay, 31-XII-1929, Stuckert 23459 (CORD). Dpto. Pilcaniyeu, a 21 km de Pilcaniyeu, en el camino a Paso Flores, 27-I-1944, Nicora 3668 (SI); Cerro Villegas, 22-XI-2000, Calviño & al. 106 (SI); RN 23 de Bariloche a Pilcaniyeu, Estancia San Ramón, junto a la ruta, 1021 m, 5-III-2008, Calviño & al. 700 y 702 (BCRU); Dina Huapi, 21-XII-1995, Puntieri 257 (BCRU); La Fragua, roquedal frente a Laguna Los Juncos, 8-XII-1992, Puntieri s.n. (BCRU); Cerro Leones, al E de San Carlos de Bariloche, XII-1966, Burkart 6221 (BA); Comallo, 7-XII-1938, Cabrera 4819 (LP). Dpto. San Antonio Oeste, RN 3 a 1 km al S de Sierra Grande, a 800 m de la ruta a Villa Hipasán, 16-XII-1993, Elechosa & al. s.n. (BAB); 10 km al sur de Sierra Grande, desvío a Punta Colorada, 05-I-1995, Chiapella & Puntieri 13 (BCRU). San Juan: Dpto. Calingasta, Los Morillos, Reserva privada de la Fundación Vida Silvestre, 2680 m, 24-I-1997, Fortunato 5587 (SI); S de Barreal, Manantiales, 3000-3300 m, 1-III-1992, Kiesling & al. 8068 (BA, MERL); Pampa Negra, 1-II-1950, Roig 3029 (LIL); entre Ojo de Agua y El Molle, 2300-2850 m, 15-XII-1976, Luti & al. 5743 (CORD). San Luis: Dpto. General Pedernera, Sierra del Morro, V-1913, Pastore 68 (SI). Santa Cruz: Dpto. Corpen Aike, Comandante Piedrabuena, 19-I-1977, Lotti de Stange 133 (LIL); 30 km al S de Comandante Piedrabuena, 19-I-1977, Lotti de Stange 139 (LIL); RN 3, Estancia Monte León, 188 m, 28-I-2003, Cocucci & Sérsic 2482 (CORD). Dpto. Deseado, 8 km al O de cruce de RP 14 y RP 68, camino hacia Bahía Mazaredo, 103 m, 22-I-2012, Fernández 24 (BCRU); camino entre Tellier y Paso Gobernador Gregores, 15-XII-1979, Bernardello & Figueroa 312, 313 (CORD); Cañadón Torcido, 64 m, 23-I-12, Fernández 25 (BCRU); Colonia General Las Heras, 19-I-1914, Ferrando 30 (SI). Dpto. Lago Argentino, Lago Viedma, 10-XII-1965, Ruiz Leal & al. 24172 bis (MERL, UC). Dpto. Lago Buenos Aires, camino de Las Heras al Lago Buenos Aires, Estancia La Porteña, 11-II-1930, Scott de Birabén 25 (LP); Río Pinturas, Grosfeld s.n. (BCRU); al S de Perito Moreno, 597 m, 11-XII-2006, Bonifacino & al. 2983 (SI); Estancia La Federica, Lago San Martin, 29-II-1936, Scott de Birabén 187 (LP); camino fronterizo, al N del Paso Rodolfo Roballos, rumbo a Los Antiguos, 24-I-2003, Belgrano 69 (SI). Dpto. Río Chico, 4 km al E del Lago Burmeister, 1050 m, 8-XII-1980, Ruiz Leal 30780 (MERL).
CHILE. Sine loc., Haenke s.n. (HAL 26839). IV Región de Coquimbo: Pcia. Elqui, 20,2 km al N de Hurtado (Iglesia) a Vicuña, 1650 m, 19-I-1993, Stuessy & Ruiz 12775a (CONC); Cordilleras de Coquimbo, Andes de Ovalle, Cerro de Huatulame, 1890, Geisse s.n. (SGO 000003751); Río Grande, XI-1890, Geisse s.n. (SGO 000003753). Pcia. Limarí, planta eléctrica Los Molles, bocatoma, 2500 m, 8-II-1963, Ricardi & al. 758 (CONC). V Región de Valparaíso: Pcia. Los Andes, cumbre del Cerro El Roble, 2200 m, 30-IV-67, Garaventa 4361 (SI); Juncal, 15 km antes de Caracoles, ruta a Mendoza, 19-I-1975, Fasanella 8340 (MERL); Uspallata-Pass der chilenischen Hochcordillere (33ºS Breite), Juncal auf den Bergen, 2300 m, 10-XI-1903, Buchtien s.n. (SI 10263); camino internacional de Los Andes a Argentina, Juncal, 2200 m, 16-I-1964, Marticorena & Matthei 629 (CONC). Pcia. Quillota, Cerro Caquis, 15 km al E de Melón 1700-2000 m, 15-XII-1938, Morrison 16915 (SI, UC); rocky summit of La Campana de Quillota, 2000 m, 28-II-1954, Constance & al. 3544 (UC). Pcia. San Felipe de Aconcagua, Andes above Río Blanco 10 km E of Río Blanco on the Trans-Andean Highway 2000-3000 m, 19-II-1954, Costance 3516 (UC). VI Región del Libertador General Bernardo O’Higgins: Pcia. Colchagua, San Fernando, Baños del Flaco, 13-XII-1937, Perez Moreau 23311 (BA); 1800 m, 7-II-1955, Ricardi 3168 (CONC); -I-1965, 1700 m, Montero 7080 (CONC); 14-I-1968, 1800 m, Montero 8082 (CONC); 30-XII-1979, 1700 m, Montero 11308 (CONC); sector huellas de dinosaurios, hacia la cumbre, 28-XII-1994, 2000-2450 m, Baeza 268 (CONC); Junta del río Tinguiririca y río del Azufre, 1220 m, 9-III-1979, Villagrán & Arroyo s.n. (CONC 55941, CONC 55947); Cajón de las Damas, 1800 m, XII-1936, Milner 22426 (CONC). Región Metropolitana de Santiago: Pcia. Santiago, San Ramón, sobre Río San Francisco, 9 km al S de Pérez Caldera, 1800 m, 12-II-1954, Constance 3506 y 3507 (CONC, UC); San Ramón, 3900 m, 5-IV-1959, Schlegel 2476 (CONC); Santuario de la Naturaleza Yerba Loca, ladera al NO del estero, sector Quebrada Agua Blanca, 2550 m, 29-II-2000, Arroyo & al. 202079 (CONC). VII Región del Maule: Pcia. Curicó, Lago Teno a Paso Vergara, 1300 m, 10-III-1967, Marticorena & Matthei 937 (CONC); Cordillera del Planchón y Portillo, Née s.n. (CONC 29919, F 68085, MA 310354); Pcia. Talca, por RN 115, aprox. a 3 km de la Laguna de Maule viniendo desde San Clemente, 2171 m, 09-II-2007, Chiapella 1667 (SI); valle del Río Maule, Río Cipreses, Laguna de la Invernada, 1300 m, 18-XII-1990, Leuenberger 4060 (CONC). VIII Región del Biobío: Pcia. Biobío, camino a Laguna El Barco, 1300 m, II-2002, Mihoc & al. 4590 (CONC); camino Estación Yumbel a Salto del Laja, 29-XII-1946, Pfister 7132 (CONC); Volcán Antuco, I-1969, Fabris & Crisci 7595 (LP). Pcia. Ñuble, Baños de Chillán, 26-II-1862, Philippi s.n. (SGO 41613); lomas sobre la fumarola, 1900 m, 16-I-1945, Pfister 946 (CONC); Cordillera de Chillán, II-1888, Philippi s.n. (K 000529783); Reserva Nacional Ñuble, 2100 m, 15-II-2003, Mihoc & al. 6921 (CONC). IX Región de la Araucanía: Pcia. Malleco, 2 km al NO de Liucura, 1000 m, 24-III-1954, Constance & Sparre 3577 (CONC, UC); Volcán Lonquimay, 1700 m, 25-III-1954, Constance & Sparre 3579 (CONC, SI, UC). XI Región de Aysén: Pcia. Aysén, Chile Chico, 230 m, 12-XII-1954, Pfister s.n. (CONC 18487). XII Región de Magallanes y de la Antártica Chilena: Pcia. Última Esperanza, cerca del Lago Pehoe, Cerro Mirador, 22-II-1986, Pedersen 14469 (SI); camino al Parque Nacional Torres del Paine, 7,2 km del camino a Cerro Guido, cerca del extremo E del Lago Sarmiento, 100 m, 24-XII-1994, Landrum & Landrum 8410 (CONC); Cerro Guido, 800 m, 16-I-1952, Pfister & Ricardi 417 (LIL).
7b. M. spinosum var. hirsutum (Phil.) Mar. Fernández & C.I. Calviño, comb. & stat. nov. M. hirsutum Phil., Anales Univ. Chile 85: 708 (1894), basónimo. TIPO: [Chile, VI Región Libertador General Bernardo O’Higgins, provincia de Colchagua:] Cordillera de San Fernando, I-1884, Hirth s.n. (lectotipo designado aquí: SGO 000003746!; isolectotipo: SGO 000003747!). Fig. 8m.
Hojas pubescentes, con pocos pelos muy largos y finos distribuidos irregularmente por toda la hoja, segmentos foliares no surcados en la cara adaxial, los laterales bipartidos —raro enteros.
Distribución geográfica.—Se distribuye por Chile: VI Libertador General Bernardo O’Higgins (Fig. 2). 1300-1800 m s.n.m.
Observaciones.—La cantidad de pelos en las hojas de M. spinosum var. hirsutum es variable y, así como se hallan ejemplares muy poco pubescentes —v.gr., Ricardi 3164 (CONC)—, también se encuentran otros con más pubescencia —v.gr., Pérez Moreau 23314 (BA) y Marticorena & Matthei 761 (CONC)—. Pese a esto, los pelos nunca llegan a cubrir las hojas completamente como en M. spinosum var. zechii.
Se designa aquí el pliego recolectado por Hirth s.n., SGO 000003746, como lectotipo entre el material tipo —F 0BN003451, K 000529782 y SGO 000003747—, dado que es el pliego con material más abundante, mejor conservado y que concuerda adecuadamente con el protólogo.
Material estudiado.—CHILE. Sine loc., 1904, Philippi s.n. (BM 001042508). VI Región del Libertador General Bernardo O’Higgins: Pcia. Colchagua, Cordillera de San Fernando, II-1888, Philippi s.n. (K 000529782); 22-II-1919, Philippi s.n. (F 0BN003451); Baños del Flaco, 14-XII-1937, Perez Moreau 23314 (BA); km 59, 1300 m, 18-I-1964, Marticorena & Matthei 761 (CONC); 1800 m, 7-II-1955, Ricardi 3164 (CONC).
7c. M. spinosum var. zechii Mar. Fernández & C.I. Calviño, var. nov. TIPO: [Chile:] III Región de Atacama: Huasco, Quebrada Yerba Buena, entre Portezuelo Yerba Buena y Río de Valeriano, 2200-3950 m, 27-I-1983, Marticorena & al. 83613 (holotipo: CONC!). Fig. 8k, l.
Hojas pubescentes, con abundantes pelos largos y finos en toda la hoja que le confieren una coloración grisácea característica; segmentos foliares surcados en la cara adaxial, con margen entero. (English: Leaves pubescent, with abundant long and fine hairs on the entire leaf, giving it a characteristic grayish color; foliar segments adaxially furrowed, with entire margin.)
Distribución geográfica.—Se distribuye por Chile: III Atacama (Fig. 2). 1000-3950 m s.n.m.
Observaciones.—Esta variedad y M. crassifolium son los únicos táxones de Mulinum que crecen en la III Región de Atacama de Chile.
Zech (1992) consideró el ejemplar de Marticorena & al. 83613 una variedad de M. spinosum que llamó var. atacamense. Sin embargo, este nombre nunca fue válidamente publicado. Dedicamos esta variedad a J.C. Zech, quien trabajó durante su formación doctoral en la sistemática del género Mulinum.
Material estudiado.—CHILE. III Región de Atacama: Pcia. Huasco, Laguna Chica, 1021 m, 17-I-1924, Werdermann 268 (SI, UC).
8. M. triacanthum Griseb., Abh. Königl. Ges. Wiss. Göttingen 19: 154 (1874). M. spinosum var. triacanthum (Griseb.) Kuntze, Revis. Gen. Pl. 3(3): 114 (1898). TIPO: [Argentina:] Catamarca in alpinis Vayas altas, Alpes de Belén, I-1872, Lorentz 586 (holotipo: GOET 008422 foto!; isotipo: CORD 00006018!). Fig. 9a-f.
|
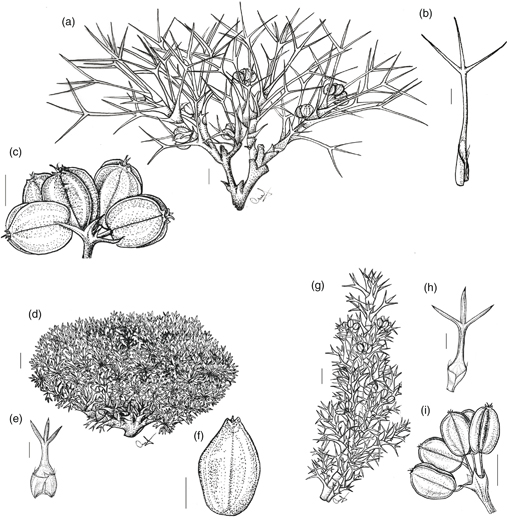 Fig. 9. Mulinum triacanthum var. triacanthum: a, fragmento de la mata; b, hoja; c, umbela con frutos. M. triacanthum var. famatinense: d, fragmento del cojín; e, hoja; f, fruto. M. ulicinum: g, fragmento de la mata h, hoja; i, umbela con frutos [a-c, Cabrera & al. 27458 (SI); d-f, Calviño & al. 817bis (BCRU); g-i, Ruiz Leal & Roig 23635 (MERL)]. [Escala: a, d, g = 1 cm; b, c, e, h, i = 2,5 mm; f = 1 mm.] Fig. 9. Mulinum triacanthum var. triacanthum: a, fragmento de la mata; b, hoja; c, umbela con frutos. M. triacanthum var. famatinense: d, fragmento del cojín; e, hoja; f, fruto. M. ulicinum: g, fragmento de la mata h, hoja; i, umbela con frutos [a-c, Cabrera & al. 27458 (SI); d-f, Calviño & al. 817bis (BCRU); g-i, Ruiz Leal & Roig 23635 (MERL)]. [Escala: a, d, g = 1 cm; b, c, e, h, i = 2,5 mm; f = 1 mm.]
|
|
Subarbusto androdioico que forma matas de 5-40 cm de altura y 10-30 cm de diámetro, bajas, irregulares, laxas, o cojines chatos, compactos. Hojas (10)11-34(39) mm, pecioladas, obovadas, rígidas; lámina 4,5-15(18) × (7)7,5-20(22) mm, trisecta, con el segmento central de 4-14,5(17) × 1-1,3(1,4) mm; segmentos de angostamente ovados a ovados o subulados, cilíndricos, a veces poco surcados en la cara adaxial, glabros, con ápice agudo, espinoso, punzante, con margen entero; pecíolo 3-14(18) mm, semejante a los segmentos, glabro; base foliar amplexicaule, con margen escasamente ciliado (raro sin cilios). Umbelas muy por debajo del nivel de las hojas, con (4)5-8 flores perfectas o (11)12-19(20) flores estaminadas; pedúnculos (3)4-6(8) mm, glabros; involucro de (3)4-5(6) brácteas de (3)4-6(7) mm, adpresas, connadas solo en la base en forma de cúpula poco profunda con lóbulos triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de (2)4-6 mm, glabros; pétalos 1,8-2 × 1-2 mm, ovados; estilopodio de algo cónico a deprimido y estilos 1-2 mm. Fruto 3-6 × 3-5(6) mm, amarillento, de contorno elíptico a anchamente elíptico, poco o muy comprimido dorsalmente; mericarpio de 1-2 mm de espesor; alas de 0,5-2(2,5) mm de anchura, de la misma anchura, más angostas o más anchas que el cuerpo del fruto, paralelas o levemente divergentes; carpóforo libre, entero o cortamente bífido.
Distribución geográfica, hábitat y fenología.—Distribuido por el noroeste de Argentina —provincias de Jujuy, Salta, Catamarca, Tucumán y La Rioja— y el sur de Bolivia —departamentos de Cochabamba, Oruro y Tarija— (Fig. 2). Abras planas de macizos montañosos altos, en pedregales y roquedales; 2200-4200 m s.n.m. Florece de enero a marzo y fructifica de mediados de enero a abril.
Observaciones.—Esta es la única especie de Mulinum que llega hasta Bolivia. Se amplía en este trabajo el área de distribución de este taxon en Argentina a la provincia de Salta.
Mulinum triacanthum es morfológicamente afín a M. ulicinum. Sin embargo, M. ulicinum tiene hojas de menor tamaño —13-17 mm—, segmentos foliares de angostamente triangulares a triangulares, más anchos —1,2-1,6 mm—, y frutos más angostos —3-4 mm— que M. triacanthum var. triacanthum. Los rangos de estos caracteres en M. ulicinum se superponen con los de M. triacanthum var. famatinense pero sus distribuciones no coinciden, ya que M. ulicinum no vive en las Sierras de Famatina, área de endemismo de M. triacanthum var. famatinense. Además, M. ulicinum se diferencia de M. triacanthum var. famatinense porque forma matas hasta de 30 cm de altura, bajas, irregulares y laxas, los segmentos foliares son triangulares y el fruto está poco comprimido dorsalmente, mientras que M. triacanthum var. famatinense forma cojines hasta de 15 cm de altura, chatos y compactos, los segmentos foliares son ovados y el fruto está muy comprimido dorsalmente.
Entre el material tipo de M. triacanthum, el pliego recolectado por Lorentz 586, GOET 008422, es el holotipo y CORD 00006018 su duplicado, tal como puede corroborarse a partir de las observaciones de Hunziker (1960) sobre los tipos grisebachianos. Allí consta que el pliego de GOET fue enviado como holotipo al Dr. Peréz Moreau y luego se constató la presencia de un duplicado en CORD.
Clave para determinar las variedades de M. triacanthumTOP
| 1. |
Matas bajas, irregulares y laxas hasta de 40 cm de altura; hojas (21)25-34(39) × (13)16-20(22) mm; segmentos foliares subulados; pecíolo (7)8-14(18) mm; fruto de contorno anchamente elíptico, poco comprimido dorsalmente, de 5-6 × 4-5(6) mm
|
8a. var. triacanthum |
| 1. |
Cojines chatos y compactos hasta de 15 cm de altura; hojas (10)11-14,5(16) × (7)7,5-9,5(10) mm; segmentos foliares de ovados a angostamente ovados; pecíolo 3-4,5(5) mm; fruto de contorno elíptico, muy comprimido dorsalmente, de 3-4 × 3-4 mm
|
8b. var. famatinense |
8a. M. triacanthum var. triacanthum. Fig. 8a-c.
M. axilliflorum Griseb., Abh. Königl. Ges. Wiss. Göttingen 19: 154 (1874), syn. nov. TIPO: [Argentina:] Catamarca en altivalles arenosos entre Nacimientos y la Laguna Blanca, 2700-3000 m, I-1872, Lorentz 430 (holotipo: GOET 008423 foto!; isotipo: CORD 00006019!).
Subarbusto que forma matas de 10-40 cm de altura y 15-30 cm de diámetro, bajas, irregulares, laxas. Hojas (21)25-34(39) mm; lámina (10)11-15(18) × (13)16-20(22) mm; segmentos subulados, no surcados en la cara adaxial, el central de (9)10-14,5(17) × (0,8)1-1,2 mm, con pecíolo de (7)8-14(18) mm. Fruto 5-6 × 4-5(6) mm, de contorno anchamente elíptico, poco comprimido dorsalmente; alas de 1-2(2,5) mm de anchura, de la misma anchura o más anchas que el cuerpo del fruto, levemente divergentes.
Distribución, hábitat y fenología.—Distribuido por el noroeste de Argentina —provincias de Jujuy, Salta, Catamarca, Tucumán y La Rioja— y Bolivia —departamentos de Cochabamba, Oruro y Tarija—. 2200-4200 m s.n.m.
Observaciones.—Se sinonimizan M. axilliflorum bajo M. triacanthum var. triacanthum, dado que no se encontraron características morfológicas ni de distribución geográfica que permitan diferenciarlos.
El pliego recolectado por Lorentz 430, GOET 008423, es el holotipo de M. axilliflorum, y CORD 00006019 su isotipo por las mismas razones explicadas para M. triacanthum (Hunziker, 1960).
Material estudiado.—ARGENTINA. Sine loc., Lorentz 136 (F 0BN003454); Cordillera de los Andes, 4000 m, 17-I-1930, Venturi 10279 (LIL). Catamarca: Sine loc., XI-1877, Lorentz s.n. (K 000529780). Dpto. Andalgalá, Andalgalá, 8-III-1916, Jörgensen 1692 (LIL, UC); Cuesta del Atajo, Cerro de las Capillitas, I-1874, Schickendantz 334 (CORD); subida a Minas Capillitas, casi llegando a la hostería Refugio del Minero, 3037 m, 13-II-2013, Fernández 57 (BCRU); subida al Globo, 3600 m, 8-XII-2010, Zuloaga 11928 (SI). Dpto. Belén, Cuesta de Belén, 2200 m, 13-II-2013, Fernández 60 (BCRU); al pie de la cuesta de Randolfo, 3200 m, 23-II-1981, Cabrera 32468 (SI); Reserva Laguna Blanca, 17-X-2002, Borgnia 32 (LP); faldeos al N del Portezuelo del Blanco, arriba de Granadillas, 3400-3500 m, 29-I-1952, Sleumer & Vervoorst 2578 (UC); entre Nacimientos y Laguna Blanca, 1872, Lorentz s.n. (BA); Vayas altas, 1872, Lorentz s.n. (SI 000739). Dpto. Tinogasta, Nevados Ojo del Salado, 15-I-1974, Grau s.n. (LIL 519837). Jujuy: Dpto. Cochinoca, cantera Huancar, cerro 5 km al S de Abra Pampa, 3500 m, 8-XI-1991, Novara & Charpin 10434 (MCNS); 3400 m, 25-XI-1986, Charpin & Ahumada 20673 (UC); Abra Pampa, II-1897, Gerling s.n. (LP 023376); 3500 m, 8-I-1972, Ruthsatz 241 (UC); Cerro Huancar, 23-I-1948, Cabrera 9431 (BAB, LP); Laguna de Guayatayoc, 3460 m, 31-XII-1971, Ruthsatz 166 (UC). Dpto. Humahuaca, Esquinas Blancas, 22-I-1966, Cabrera & al. 17734 (UC); 3650 m, 5-II-1971, Ruthsatz 35 (BAA, UC); paradero ferrocarril km 1321, junto a la RN 9, 7-II-1979, Arenas 947 (SI); Tres Cruces, 21-I-1976, Cabrera 27458 (SI); Cerro Tres Cruces enroute to Tres Cruces, 3700 m, 11-I-1968, Costance & Cabrera 3854 (UC); Cabrera & al 18947 (UC); Azul Pampa, 3350 m, 15-II-1901, Claren 11702 (CORD). Dpto. Yavi, Cangrejillos, 3600 m, 13-II-1972, Ruthsatz 363 (UC). La Rioja: Dpto. Chilecito, sine loc., II-1905, Stuckert 15125 (CORD, UC); Cerro Negro, 3650 m, 10-I-1908, Salazar 15209 (CORD); Sierra de Sañogasta, falda O, subiendo desde Aicuña hacia el ESE, rumbo al Cerro Homónimo, 2300 m, 17-XII-1975, Hunziker 22843 (CORD). Dpto. Famatina, Sierra de Famatina, Famatina, Barboza & Cantero 3932 (CORD); Río Amarillo, 3650 m, 4-II-1995, Ruthsatz 9199 (UC); Quebrada de la Cunsche, 3500 m, 2-IV-1949, Krapovickas 6250 (BAB, UC); Tres Piedras, 3285 m, 3-II-2011, Barboza & al. 2925 (CORD); entre la bifurcación hacia Tres Piedras y Los Tambitos; 3458 m, 3-II-2011, Barboza & al. 2897 (CORD); camino a la Mina La Mejicana, pasando Los Berros, 3056 m, 5-XI-2013, Calviño & al. 808 (BCRU); 3106 m, 5-XI-2013, Calviño & al. 809 (BCRU); 3290 m, 5-XI-2013, Calviño & al. 813 (BCRU); 3502 m, 5-XI-2013, Calviño & al. 814 (BCRU); Cueva de Pérez, 3846 m, 5-XI-2013, Calviño & al. 816 (BCRU); desde Cueva de Pérez a la Mejicana, 4032 m, 5-XI-2013, Calviño & al. 818 (BCRU); entre Corrales y Cueva de Pérez, 3800 m, 13-I-1976, Cabrera & al. 27181 (SI); 2 km antes de llegar a la Cueva de Pérez, 3229 m, 9-II-2013, Fernández 50 y 51 (BCRU); bajando de la Mina La Mejicana, 4014 m, 1-II-2011, Barboza & al. 2576 (CORD); Ciénaga de Cosme, 19-I-1928, Castellanos s.n. (BA 28/291); pueblo de Santa Rosa, 3100 m, 14-I-1947, Hunziker 1884 (SI); Cueva de Medina, 3100 m, 31-III-1949, Krapovickas 6184 (BAB, CORD); Quebrada del Río Peñón 3000 m, 01-II-1947, Hunziker 2099 (CORD, UC); Mina San Juan, 27-II-1906, Kurtz 13655 (CORD); entre la Mina Yareta y la altura de Espíritu Santo, 25-I-1879, Hieronymus & Niederlein 792 (CORD); entre Cerro Negro y la altura de Espíritu Santo, 26-I-1879, Hieronymus & Niederlein 819 (CORD). Dpto General La Madrid, Cerro Leoncito, en la quebrada, 10-XII-1915, Hosseus 1151 (CORD). Salta: Dpto. Cachi, Aguada de Guanaco, laguna 2 km al S de RN 33 entre Cachipampa y Tin-Tin, 3000-3300 m, 29-XII-1990, Novara & Osten 10078 (MCNS); 23 km después de carrefour de la RN 33 en dirección de Cerro Negro, 3630 m, 05-I-1994, Charpin & Lazare 24050 (UC); ca. 1 km ruta Amblayo, ruta de Mina Don Otto, 3100 m, 04-I-1994, Charpin & Lazare 24022 (UC). Dpto. San Carlos, Alturas de Amblayo, valle de Isonza, 3200 m, 25-III-1987, Novara 6347 (MCNS); Cuesta de Isonza, 20 km al S de Piedra del Molino, 3000-3500 m, 1-IV-1990, Novara & Neumann 9775 (MCNS). Tucumán: Dpto. Tafí del Valle, cumbres Calchaquíes, Quebrada del Real, 4200 m, 31-I-1907, Dinelli 542 (BAB); 3200 m, 30-I-1912, Rodríguez 293 (SI, UC); Cerro Muñoz, 3600 m, 10-I-1979, Halloy 583 (LIL); Mesada de los Yutos, 16-III-1962, Krapovickas 10655 (LIL).
BOLIVIA. sin. loc. 3850 m, 18-II-1980, Ruthsatz 507 (UC). Cochabamba: Prov. Tapacarí, Viscachani, 2,5 km NNW de Japa K’asa, km 125, 4120 m, 23-III-1995, Pestalozzi 474 (LPB). Oruro: Prov. Cercado, 35 km de Caracollo, Lequepalca, 4 km hacia Cochabamba, 3950 m, 15-I-1995, Beck 21726 (LPB). Tarija: Prov. Aviles, Puna Patanca, 3800 m, 7-I-1904, Fiebrig 2601 (SI).
8b. M. triacanthum var. famatinense (H. Wolff) Mar. Fernández & C.I. Calviño, comb. & stat. nov. M. famatinense H. Wolff, Repert. Spec. Nov. Regni Veg. 17: 441 (1921), basónimo. TIPO: [Argentina:] La Rioja, Sierra de Famatina, Cueva de Pérez, 26/28-I-1879, Hieronymus & Niederlein 362 (lectotipo, designado aquí: CORD!; isolectotipo: B destruido, foto F neg. 3452!). Fig. 9d-f.
Subarbusto que forma cojines de 5-15 cm de altura y 10-20 cm de diámetro, chatos, compactos. Hojas (10)11,5-14,5(16) mm; lámina 4,5-5,5(7) × (7)7,5-9,5(10) mm; segmentos de angostamente ovados a ovados, a veces poco surcados en la cara adaxial, el central de 4-7,5(8) × 1-1,3(1,4) mm; pecíolo 3-4,5(5) mm. Fruto 3-4 × 3-4 mm, de contorno elíptico, muy comprimido dorsalmente; alas de 0,5 mm de anchura, de la misma anchura o más angostas que el cuerpo del fruto, paralelas.
Distribución, hábitat y fenología.—Se distribuye por el noroeste de Argentina, en la provincia de La Rioja, endémico de la Sierra de Famatina. 4000-4700 m s.n.m.
Observaciones.—Mulinum famatinense hasta ahora era conocido solo por su tipo y ha sido tratado como especie (Martínez, 2008). Sin embargo, sobre la base de la variación morfológica asociada a la altitud que presenta M. triacanthum en la Sierra de Famatina y de estudios filogenéticos (obs. pers. y Fernández & al., 2017b), consideramos M. famatinense una variedad de M. triacanthum.
El único material que se cita en el protólogo de M. famatinense es el recolectado por Hieronymus & Niederlein 364 en la Cueva de Pérez —Sierra de Famatina—. Este pliego no ha sido encontrado en ninguno de los herbarios consultados —v.gr., B, F, CORD y en el cibersitio Plant Science en JSTOR—. En cambio, sí se localizaron pliegos de Hieronymus & Niederlein con numeración 360 y 362 que coinciden con el protológo en descripción, recolectores y localidad de recolección. Según el art. 9.12 del CINB (McNeill & al., 2012), si no hay isotipos, ni sintipos, ni isosintipos, ni paratipos, el lectotipo debe elegirse entre los ejemplares no citados o ilustraciones que comprendan el resto del material original. El pliego de B Hieronymus & Niederlein 362 fue visto por H. Wolff con anterioridad a la publicación de la especie, como consta en una etiqueta —no hay registros de Hieronymus & Niederlein 360 vistos por Wolff—. Por lo tanto, según el art. 9.3a del CINB (McNeill & al., 2012), puede considerarse al menos a la recolección numerada con el 362 como parte del material original ya que, sin duda, corresponde a ejemplares con los que Wolff hizo la descripción. Como el pliego de B está destruido y solo se dispone de una fotografía de F, se selecciona su duplicado, un pliego de CORD, como lectotipo. Este pliego, además de estar de acuerdo perfectamente con el protólogo, presenta abundante material vegetativo y frutos.
Material estudiado.—ARGENTINA. La Rioja: Dpto. Famatina, Sierra de Famatina, Cueva de Pérez, 26/28-I-1879, Hieronymus & Niederlein 360 (CORD); desde Cueva de Pérez a la Mina La Mejicana, 4032 m, 5-XI-2013, Calviño & al. 817, 817bis (BCRU); 4005 m, 5-XI-2013, Calviño & al. 819 (BCRU); Mina La Mejicana, 4326 m, 1-XII-2011, Barboza & al. 2550, 2555 (CORD); estación 8 pasando la Mina La Mejicana, 4227 m, 18-XII-2011, Barboza & al. 3163 (CORD); Real Viejo, 4078-4115 m, 20-XII-2011, Barboza & al. 3350 (CORD); camino al Alto Blanco, 20-I-1928, Castellanos s.n. (BA 28/292); Los Bayitos, 4500 m, 7-IV-1949, Krapovickas 6337 (CORD); Cerro La Fofosa (cumbre), 4682 m, 1-III-2013, Barboza & Cantero 3925 (CORD).
9. M. ulicinum Gillies & Hook., Bot. Misc. 1: 328, lám. 64 (1830). TIPO: [Argentina, Mendoza:] Montañas de Uspallata, Gillies s.n. (lectotipo, designado aquí: K 000529781 foto!; isolectotipos: E 00000044 foto!, E 00000045 foto!). Fig. 9g-i.
M. triacanthum var. multiflorum Hieron., Bol. Acad. Ci. (Córdoba) 4: 27 (1881), syn. nov. M. ulicinum var. multiflorum (Hieron.) Zech, Kurtziana 24: 192 (1995). TIPO: [Argentina:] San Juan, Leoncito, II-1876, Echegaray s.n. (holotipo: CORD 00006705!).
Subarbusto androdioico que forma matas de 5-30 cm de altura y 15-30 cm de diámetro, bajas, irregulares, laxas, resinosas. Hojas (11)13-17(19) mm, pecioladas, obovadas, rígidas; lámina (4)5-7(8) × (5)6-10(11) mm, trisecta, con el segmento central de 4-6(7) × (1) 1,2-1,6(2) mm; segmentos de angostamente triangulares a triangulares, cilíndricos, surcados en la cara adaxial, glabros, con ápice agudo, espinoso, punzante, con margen entero; pecíolo (2)3-5(7) mm, semejante a los segmentos, glabro; base foliar amplexicaule, margen escasamente ciliado —raro sin cilios—. Umbelas muy por debajo del nivel de las hojas, con (2)4-6 flores perfectas o 7-12(13) flores estaminadas; pedúnculos 3-5(7) mm, glabros; involucro de (3)4-5(6) brácteas de (2)3-3,5(4) mm, adpresas, connadas casi completamente en forma de cúpula profunda con lóbulos anchamente triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 2-4(5) mm, glabros; pétalos 1,5-2 × 1-1,8 mm, ovados; estilopodio de algo cónico a deprimido y estilos 1-1,2 mm. Fruto (3)4-5 × (2)3-4 mm, amarillento, de contorno elíptico, poco comprimido dorsalmente; mericarpio de 1-1,5 mm de espesor; alas de (0,5)1 mm de anchura, más angostas que el cuerpo del fruto, paralelas; carpóforo libre, entero o cortamente bífido.
Distribución geográfica, hábitat y fenología.—Se distribuye por el noroeste y el oeste de Argentina —provincias de Jujuy, Salta, Catamarca, Tucumán, La Rioja, San Juan y Mendoza— y el norte de Chile —II Antofagasta— (Fig. 2). Abras planas y expuestas, entre rocas; 2700-4900 m s.n.m. Florece de diciembre a marzo y fructifica de diciembre a abril.
Observaciones.—Se amplía en este trabajo el área de distribución de esta especie a las provincias de Catamarca, Tucumán y La Rioja en Argentina. Mulinum ulicinum fue descrita para Argentina y su presencia en Chile se consideraba dudosa (Martínez, 2008). Sin embargo, la hemos recolectado tanto en el lado argentino como en el chileno a la altura del Paso de Jama, que une Jujuy con la II Región de Antofagasta. De hecho, M. ulicinum había sido ampliamente recolectada en Chile, aunque se encontraba mal determinada, como M. crassifolium, en los herbarios (Fernández & al., 2014). En los tratamientos realizados por Clos (1848-1849), Weddell (1857) y Reiche (1899) sobre la flora de Chile, se incluye M. ulicinum. Sin embargo, tal como se detalla en Fernández & al. (2014), estas citas de M. ulicinum para Chile se debieron a confusiones con otras especies.
Mulinum ulicinum es morfológica y filogenéticamente afín a M. triacanthum (véanse las observaciones hechas para esta especie). Además, es fácil de confundir con Azorella cryptantha (Clos) Reiche, debido a sus frutos inconspicuamente alados y hojas cilíndricas, rígidas y punzantes. Sin embargo, Azorella cryptantha forma cojines chatos que no superan los 8 cm de altura y tiene pecíolos muy largos —hasta de 20 mm— (Martínez, 1989) en comparación con los de M. ulicinum.
Mulinum triacanthum var. multiflorum es ubicada por Martínez (2008) en la lista de los sinónimos de M. triacanthum. Sin embargo, a partir del estudio de su material tipo y de análisis morfométricos (obs. pers.), no se han observado diferencias morfológicas entre esta entidad y el material tipo y ejemplares de herbario de M. ulicinum, por lo que se considera sinónimo de M. ulicinum.
La fotografía del material tipo de M. ulicinum tiene un parecido morfológico con lo fotografiado del material tipo de M. echegarayi. Sin embargo, M. echegarayi se diferencia claramente de M. ulicinum por el menor tamaño de las hojas —8-10 mm— y los segmentos planos, con ápice de obtuso a redondeado y mucronado en M. echegarayi.
Se designa el pliego recolectado por Gillies s.n., K 000529781, como lectotipo de M. ulicinum entre sus duplicados, por contener la mayor cantidad de material.
Material estudiado.—ARGENTINA. Catamarca: Dpto. Andalgalá, Río Potrero sup., La Overa, 4100 m, 1-III-1951, Sleumer 1874 (LIL). Dpto. Antofagasta de la Sierra, vega, 4200 m, Rodríguez 32 (SI). Dpto. Belén, Valle del Río Agua Calientes, 4200 m, 23-I-1995, Martínez Carretero 1122 (MERL). Dpto. Tinogasta, paso San Francisco, sin col. (LIL 64433). Jujuy: Dpto. Humahuaca, Mina Aguilar, 4000 m, 11-I-1968, Cabrera & al. 18964 (UC); 4300 m, 12-I-1968, Constance 3856 (SI, UC); 4400 m, 14-I-1948, Cabrera 9221 (LP); 14-I-1948, Cabrera 9214 (LP); molino, 4300 m, 3-II-1972, Ruthsatz 336 (BAA, UC); veta, 4500 m, 27-I-1971, Ruthsatz 7 (BAA, UC); toma de agua, 4950 m, 20-I-1971, Ruthsatz 41 (BAA, UC); toma de agua de molino, 11-II-1971, Ruthsatz 57 (BAA, UC); espinazo del diablo, 3800 m, 19-I-1972, Ruthsatz 275 (BAA, UC); 7-II-1971, Ruthsatz 40 (BAA, UC); 3800 m, 17-I-1971, Ruthsatz 47 (BAA, UC); bajada al Río Despensa, 4000 m, 13-II-1970, Ancibor & Mujica 32 (BAA, UC); Río Despensa, 4100 m, 31-I-1971, Ruthsatz 44 (BAA, UC); Cerro arriba del molino, 4100 m, 16-I-1953, Sleumer 3365 (LIL); camino al campamento viejo (camino a la veta), 4400 m, 16-XII-2010, Calviño & al. 797 (BCRU); arriba de la mina cerca de toma de agua, 4670-4730 m, 4-III-1983, Hunziker 10605 (SI). Dpto. Rinconada, alrededores de Laguna Vilama, 4600 m, 5/12-III-1964, Schwabe & al. 1037 (LP); Campo Lagunilla (camino hacia Aparzo), 4100 m, 7-II-1979, Gonzalez 22 (LIL). Dpto. San Antonio de los Cobres, Quebrada de Polvorillas, 4200 m, 2-II-1944, Cabrera 8353 (LP). Dpto. Susques, Sey, 12-II-1946, Cabrera 9069 (LP, UC); 4100 m, 2-I-1972, Ruthsatz 204 (UC); unos kms después del mirador del salar de Jama, 4250 m, 14-XII-2010, Calviño & al. 793 (BCRU); Abra de Chorrillos, 4500 m, 16-XI-1980, Cabrera 31775 (SI); 2-I-1972, Ruthsatz 210 (UC); Cueva Colorada próximo a Achibarca, 12-III-1927, Castellanos s.n. (BA 27/846); al norte de la Salina Olaroz, 4080 m, 3-I-1972, Ruthsatz 225 (UC). Dpto. Tumbaya, Alto del Morado, entre desvío a Tres Morros y Purmamarca, 4200 m, 15-XII-2010, Calviño & al. 794 (BCRU); arriba de San Antonio (Abra) de Los Pibes, 4250 m, 7-I-1953, Sleumer 3311 (LIL, SI); Abra de Los Pibes, 4000 m, 17-XII-1971, Ruthsatz 126 (BAA, UC); al E del Abra de Los Pibes, 3870 m, 18-XII-1971, Ruthsatz 136 (BAA, UC); 3980 m, 18-XII-1971, Ruthsatz 135 (BAA, UC); El Moreno, 3-II-1901, Fries 840 (CORD); Volcán, II-1920, Castillón 7188 (LIL). Dpto. Valle Grande, Caspalá, cumbres, 1-III-1940, Burkart & Troncoso 11811 (SI). La Rioja: La Rioja, Giacanelli s.n. (LIL 64434). Dpto. Sarmiento, Quebrada del Río Salado, cercanía al Cerro Bonete, 4400 m, 2-III-1950, Hunziker 4145 (BAB, CORD, UC). Mendoza: Dpto. Las Heras, Alto de los Manantiales, 3030 m, 19-II-1965, Ruiz Leal & Roig 23635 (MERL); 2900-3100 m, 19-II-1965, Roig 5274 (MERL); 20-II-1965, Roig 5234 (MERL) 2830 m, 21-II-1965, Ruiz Leal & Roig 23676 (MERL); 19-II-1965, Roig 5274 (MERL); Los Vallecitos, 2700 m, 6-I-1951, Novara 13400 (MERL, UC). Dpto. Malargüe, Malargüe, entre Arroyo Chaguitracó y Portezuelo del Choique, 16-I-1958, Ruiz Leal & Roig 18833 (MERL, UC). Dpto. Luján de Cuyo, zona del Cerro del Plata, Piedra Grande, 3600-3700 m, 12-I-1960, Ruiz Leal & al. 21045 (MERL). Dpto. San Carlos, Estancia Yaucha, Pampa de las Osamentas, 3000 m, 18-I-1941, Ruiz Leal & al. 7182 (MERL, UC). Dpto. Tunuyán, Tunuyán, 20-III-1935, Ruiz Leal & al. 3184 (MERL nacimiento del Arroyo de las Cuevas, 3200 m, 23-III-1938, Norman 3126 (BA, MERL, UC); Paso del Portillo, Cuesta de los Afligidos, 29-I-1934, Ruiz Leal & al. 2034 (BA, MERL); 24-XII-1933, Ruiz Leal & al. 1855 (BA, LIL, MERL, UC). Salta: Dpto. Cafayate, Sierras de Cajón, 4500 m, 6-III-1944, Rodríguez 1338 (LP). Dpto. Coronel Moldes, Mina Concordia, Gobernación de los Andes, 4500 m, 01-X-1929, Mintzer 60101 y 60102 (BAB). Dpto. La Poma, Abra de Acay, RN 40, 4750 m, 23-I-1989, Arriaga & al. 426 (BA); 4895 m, 18-II-1986, Varela & Del Castillo 1362 (MCNS); faldeo NO del Abra de Acay, expedición al Llullaillaco, 4200 m, Bravo s.n. (LIL 539722); Cerro Tuzgle, cabecera del volcán, 4600-4800 m, 4-III-1993, Nuñez 763 (MCNS); 4200 m, 2-II-1944, Cabrera 8378 (LP). Dpto. Los Andes, camino a Alto de Tocomar, km 181 de la RN 51, 4500 m, 17-XII-2010, Calviño & al. 798 (BCRU); 4300 m, 27-II-1972, Cabrera & al. 22540 (UC), Abra del Gallo, 4600-4650 m, 4-III-1993, Nuñez 751 (MCNS); 4750 m, 12-II-1945, Cabrera 8672 (LP); aprox. 30 km al SO de San Antonio de los Cobres en el camino de herradura de Pastos Grandes, 17-XII-1946, Krapovickas 3212 (LIL); San Antonio de los Cobres 27 km hacia paso Huaitiquina, abra Chorillos, 4560 m, 30-XII-1986, Beck 14141 (UC). San Juan: Dpto. Calingasta, Río Manrique a Portezuelo de Potrerillos, 3000 m, 23-I-1991, Kiesling 7688 (SI); SO de Barreal, zona de El Mercedario, cordón de la Ramada, 3200 m, 30-XII-2010, Kiesling 10408 (BCRU, MERL); Espinacito, Río de las Leñas, 3600 m, 1-II-1949, Roig 11949 (MERL, UC); Paso del Espinacito, 2-I-1953, Castellanos s.n. (LIL 15501); Paso del Espinacito a Los Hornillos, 23-I-1953, Castellanos s.n. (LIL 420516); Ao. de la Ramada, 19-I-1951, Semper s.n. (MERL 45540); 4200 m, 9/21-I-1951, Semper 13959 (LIL). Dpto. Iglesia, Valle del Cura, Cerro Conconta, 4000 m, 24-I-1981, Kiesling 3228 (SI); Cordillera de Colangüil: de Invernadas a la Quebrada del Salto, 15-I-1930, Perez Moreau s.n. (BA 30/147); Rodeo-El Pierro, Arroyo Los Ocucaros, 3460 m, 27-I-1995, Ruthsatz 9073 (UC). Tucumán: Dpto. Tafí del Valle, Cumbres Calchaquíes, 4600 m, 21-I-1907, Stuckert 17894 (CORD); línea de cumbre, 4260 m, 24-03-2002, Ayarda & Muruaga 173 (LIL); entre La Puerta y Los Cardones, 4200 m, 30-I-1933, Burkart 5344 (SI, UC); Cerro Santa Isabel, 4600 m, 19-II-1990, Ayarda 363 (LIL); Quebrada de Matadero, 4200 m, I-1963, De la Sota 2725 (LIL).
CHILE. II Región de Antofagasta: Pcia. El Loa, a 10 km del límite con Argentina, 4430 m, 14-XII-2010, Calviño & al. 792 (BCRU); Cerro Curutú, lado S del Paso de Jama, 4700 m, IV-1997, Arroyo & al. 97268 (CONC); cerro nevados de Poquis, ladera SO, 4500 m, IV-1997, Arroyo & al. 97385 (CONC); cordón Ceja Alta, lado SE, 4600 m, IV-1997, Arroyo & al. 97290 (CONC); borde sur del Salar de Tara, 4120 m, 18-XII-1996, Moreira & Muñoz 300 (CONC); cruce entre Paso de Jama y Salar de Tara, 4250 m, 19-XII-1996, Moreira & Muñoz 318 (CONC).
10. M. valentini Speg., Anales Soc. Ci. Argent. 48: 57 (1899). TIPO: [Argentina:] Chubut: Trelew, XI-1897, Valentín s.n. (lectotipo, designado aquí: LP 002800!; isolectotipo: SI 000740!). Fig. 10.
|
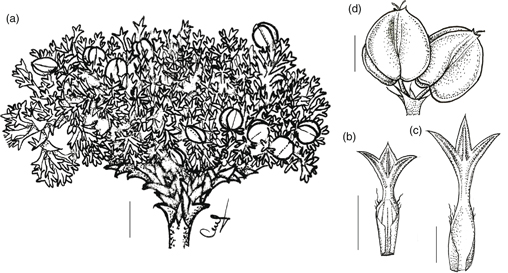 Fig. 10. Mulinum valentini: a, fragmento del cojín; b, c, hojas de distintos cojines; d, umbela con frutos [a, b, d, Ruiz Leal & al. 26434 (MERL); c, Fernández 30 (BCRU)]. [Escala: a = 1 cm; b, c = 2 mm; d = 2,5 mm.] Fig. 10. Mulinum valentini: a, fragmento del cojín; b, c, hojas de distintos cojines; d, umbela con frutos [a, b, d, Ruiz Leal & al. 26434 (MERL); c, Fernández 30 (BCRU)]. [Escala: a = 1 cm; b, c = 2 mm; d = 2,5 mm.]
|
|
M. lycopodiopsis Speg., Anal. Soc. Ci. Argent. 48: 55 (1899). TIPO: [Argentina, Santa Cruz:] Patagonia, Pan de Azúcar, Río Chico, XII-1897, Ameghino s.n. (lectotipo, designado aquí: LP 002796!; isolectotipo: BA 27292!).
Azorella concolor Rendle, J. Bot. 42: 368 (1904). TIPO: [Argentina, Santa Cruz:] Península Burmeister, Lago Argentino, I-1900, Prichard s.n. (holotipo: BM 001008565 foto!).
Subarbusto andromonoico que forma cojines de 3-10 cm de altura y 10-30 cm de diámetro, chatos, compactos. Hojas 8-13(15) mm, subsésiles, de obovadas a anchamente obovadas, de semirígidas a rígidas; lámina (2)3-5 × 4-6(7) mm, trisecta —raro tripartida—, con el segmento central de 2-4 × (1,2)1,4-2,2(2,4) mm; segmentos de triangulares a anchamente triangulares, planos, surcados en la cara adaxial, glabros, con ápice de agudo a obtuso, espinuloso, punzante, con margen entero; pecíolo 1-3(4) mm, semejante a los segmentos, glabro; base foliar amplexicaule, con margen ciliado. Umbelas que apenas sobrepasan el nivel de las hojas, con 1-3 flores perfectas o 4-7 flores estaminadas; pedúnculos (2)3-4(5) mm, glabros; involucro de 3-5 brácteas de 2-3 mm, adpresas, connadas casi completamente en forma de cúpula profunda con lóbulos angostamente triangulares, glabra en el interior, con margen entero y ápice agudo. Flores con pedicelos de 2-4 mm, glabros; pétalos 1,5-2,1 × 1-1,8 mm, ovados; estilopodio cónico y estilos 1-1,5 mm. Fruto (4)5(7) × (3)5-6(7) mm, de amarillento a rojizo, de contorno anchamente elíptico a circular, poco comprimido dorsalmente; mericarpio de 1-2,5 mm de espesor; alas de 1-3 mm de anchura, más anchas que el cuerpo del fruto, divergentes; carpóforo persistentemente unido a uno de los mericarpios.
Distribución geográfica, hábitat y fenología.—Se distribuye por el sur de la Patagonia argentina —provincias de Chubut y Santa Cruz— y el extremo sur del Chile continental —regiones XII Magallanes y de la Antártica Chilena— (Fig. 2). En terrenos arenosos entre piedras sueltas o en afloramientos rocosos; 500-1600 m s.n.m. Florece y fructifica de noviembre a marzo.
Número cromosómico.—n = 8 (Constance & al., 1971; Constance, 1988).
Nombre vernáculo.—Yareta (Argentina; Constance, 1988).
Observaciones.—El epíteto dedicado a J. Valentin debe escribirse con una sola i final, de acuerdo con la recomendación 60C.2 del CINB (McNeill & al., 2012), ya que Valentín posee una forma latinizada bien establecida (Valentinus).
Mulinum valentini se asemeja a M. albovaginatum, pero este último tiene hojas más grandes —6-8 mm—, con segmentos foliares ovados y frutos más angostos —3-4 mm—, mientras que el primero tiene hojas más grandes —8-13 mm—, segmentos foliares triangulares y frutos más anchos —5-6 mm—. Además, tienen distribuciones alopátricas y la de M. valentini es la más austral.
Se designa aquí el pliego recolectado por Valentín s.n., LP 002800, como lectotipo de M. valentini entre sus duplicados, porque se encuentra mejor conservado, presenta abundante material vegetativo y umbelas floríferas y concuerda perfectamente con el protólogo. También se designa al pliego de Ameghino s.n., LP 002796, del cerro Pan de Azúcar como lectotipo de M. lycopodiopsis, ya que presenta material más abundante y un duplicado —isolectotipo en BA—, a diferencia del sintipo del Lago Argentino —LP 002797.
Material estudiado.—ARGENTINA. Chubut: Dpto. Escalante, 70 km al norte de Comodoro Rivadavia, 640 m, 6-I-1991, Leuenberger 4118 (BAB); Pampa del Castillo, 17-XII-1981, Cabrera & al. 33250 (SI); 28-I-1932, Castellanos s.n. (BA 6119, UC); 02-II-1932, Castellanos s.n. (BA 6117, UC); RP 26 a 85 km al este de Sarmiento y a 68 km al oeste de Comodoro Rivadavia, a través del punto Panorámico (yacimientos de petróleo), 870 m, 27-I-1989, Zech & Contogiorgakis 24 (CONC, UC); campo Diadema Norte, 30 km al NW de Comodoro Rivadavia, 16-XII-1952, Ruiz Leal & al. 14784 (MERL, UC). Dpto. Languiñeo, Tecka, Estancia La Carlota, 23-I-1949, Grondona 2202 (UC). Dpto. Paso de los Indios, RN 23, a 76 km al N de Buen Pasto, 2-XII-1976, Arroyo & al. 223 (SI); top of Mt. Quichaura ca. 100 km south of Esquel, 7-IV-1952, Beetle & Soriano 509 (UC). Dpto. Río Senguer, Pampa de Chaliá, Estancia Media Luna, 3-XII-1981, Villamil & al. 2228 (SI). Dpto. Sarmiento, bosque petrificado de María Santísima a 60 km SW de Sarmiento, 4-XII-1967, Correa & al. 4090 (UC). Dpto. Tehuelches, RP 44, Laguna de los Niños, camino al Lago Vintter, 1101 m, 25-I-2003, Cocucci & Sérsic 2466 (CORD, SI); entre Río Pico y Lago Vintter, 16-I-1948, Soriano 3072 (SI, UC); ca. 3 kms E of Lago Vintter on gravel road to Río Pico, 930 m, 7-XII-1984, Stuessy & al. 6892 (SI); Estancia El Cherque, 30 km al E de Río Pico, 26-XI-1946, Soriano 2160 (UC); valle del Lago Blanco, 22-XII-1902, Koslowsky 12353 (BAB, SI, UC); Koslowsky 1314, 1318, 1319 (BA, UC); Estancia Rosales, 1000 m, 28-I-1978, Halloy 39 (LIL); Arroyo Temenhuan, 29-XI-1946, Soriano 2193 (UC). Santa Cruz: Dpto. Deseado, colonia Las Heras, Cerro Romberg, 22-I-1965, Ancibor 119 (UC). Dpto. Güer Aike, Río Gallegos, Estancia La Carlota, 9-XII-1975, Arroyo & al. 358 (BAB, UC), Estancia La Angelina, 24-I-1979, Faggi & López 224 (UC); Estancia La Verdadera Argentina, Cerro de la Virgen, 900 m, Arroyo & al. 2252 (UC); Estancia Las Vizcachas, Cerro Las Vizcachas, 25-I-1977, Arroyo & al. 382 (UC); Estancia Las Vizcachas, cono volcánico Pan de Azúcar, 1030 m, 28-I-1977, Arroyo & al. 508 (UC); ladera N, 29-I-1977, Arroyo & al. 562 (UC); Laguna Vizcachas, ladera SSE de la laguna, 1200 m, 28-I-1977, Arroyo & al. 2610 (CONC, UC); aprox. 2 km de Laguna Vizcachas, 23-I-1977, Arroyo & al. 2384 (BAB, UC); Cerro de las Vizcachas, 710-900 m, 17-I-1970, Ruiz Leal & al. 27020 (MERL, UC). Dpto. Lago Argentino, Lago Argentino, Karr-aik, III-1898, Ameghino s.n. (LP 002796); cumbre del Cerro El Calafate, 868 m, 20-XII-1950, Sleumer 1178 (LIL); Cerro Huiliche, Laberinto, Estancia Anita, 12-I-2009, Guerrido & Fernández 831 (SI). Dpto. Lago Buenos Aires, a 88 km de la localidad Perito Moreno, 13-XII-1954, Soriano 4795 (BAB, UC); RN40, 5,3 km al S del ingreso a la Estancia Cueva de las Manos, al S de Perito Moreno, 597 m, 11-XII-2006, Bonifacino & al. 2982 (SI); 64 km al S de Perito Moreno, 569 m, 25-I-12, Fernández 30 (BCRU); a 20 km de Bajo Caracoles hacia Perito Moreno, 28-I-1967, Boelcke & al. 12912 (UC); meseta del Lago Buenos aires, 1300-1500 m, 23-XII-1968, Ruiz Leal & al. 26434 (MERL, UC); meseta del Lago Buenos Aires, Laguna El Sello, 1600 m, 24-XII-1968, Ruiz Leal & al. 26451 (MERL, UC); meseta del Lago Buenos Aires, a 10 km de Estancia La Vizcacha, 1200 m, 23-XII-1968, Ruiz Leal & al. 26423 (MERL, UC); margen de la meseta del Lago Buenos Aires, arriba de la Estancia Pérez (Río de las Vueltas), 1100-1350 m, 28-XII-1950, Sleumer 1358 (LIL). Dpto. Río Chico, Lago Posadas, 750 m, 12-XII-1965, Ruiz Leal & al. 24225 (MERL, UC).
CHILE. XII Región de Magallanes y de la Antártica Chilena: Pcia. Última Esperanza, Sierra de los Baguales, Estancia La Cumbre, 1000 m, 13-XII-2000, Domínguez 286 (CONC); 1200 m, XII-1986, Landero 643 (CONC); Campo de la Tropilla, 900 m, 17-XII-1986, Landero 699 (CONC); Cerro Donoso, Rio de las Chinas, 900 m, 12-II-1987, Arroyo & al. 870276 (CONC); Cerro Santa Lucía, 1000 m, 1-XII-1984, Arroyo & al. 841002 (CONC); Sierra del Cazador, 22-II-1955, Magens 918 (CONC, UC).
Nombres dudososTOP
M. spinosum f. humilis Kurtz, Verh. Bot. Vereins Prov. Brandenburg 35: 111 (1894).
No se ha logrado encontrar el material tipo, pese a exhaustivas búsquedas, principalmente en B, F, CORD y NY, donde existen colecciones estudiadas por Kurtz.
M. manifolium Ravenna, Onira 11(21): 56-57 (2008).
Este taxon fue publicado por Ravenna a partir de un único ejemplar recolectado por él mismo en el macizo de Cantillana, en la Región Metropolitana de Chile. Según Ravenna (2008), el holotipo fue depositado en el herbario SI; sin embargo, este material nunca llegó a dicha Institución —N. Deginani, com. pers.—. Hasta que no se halle y estudie este material, consideramos este nombre dudoso.
Nombres excluidosTOP
M. acaule (Cav.) Pers., Syn. Pl. 1: 309 (1805). Selinum acaule Cav., Icon. 5: 59-60, lám. 487 fig. 2 (1799). TIPO: [Argentina, Santa Cruz:] Puerto Deseado, Née s.n. [lectotipo, designado por Fernández & al. (2016b): MA 476255 foto!) = Laretia acaulis (Cav.) Gillies & Hook., Bot. Misc. 1: 329, lám. 65 (1830).
M. angulatum DC., Prodr. 4: 80 (1830). TIPO: Chile, Cordillera, Haenke s.n. (lectotipo, designado aquí: PR 179031 foto!; isolectotipo: PRC 450111 foto!) = Pozoa coriacea Lag., Gen. Sp. Pl. 13 (1816).
Se designa el pliego PR 179031 como lectotipo de M. angulatum entre los dos duplicados por ser el que presenta mayor abundancia de material. El pliego de PR no posee etiquetas que indiquen el recolector, a diferencia del de PRC. Sin embargo, según informó el conservador de PR (Otakar Sída, com. pers.), el pliego fue recolectado por Haenke y es un duplicado del de PRC. Ambos provienen de una donación hecha al Museo de Praga luego de la muerte de Haenke. Originalmente, la colección sólo presentaba pequeñas anotaciones de Haenke con los lugares de recolección, en grupos de pliegos. Las etiquetas que se observan en los pliegos actualmente corresponden a transcripciones hechas por C. Sternberg y C. Presl y, en el caso de los pliegos de la familia Apiaceae, también por De Candolle, a quien se le enviaron a Ginebra para su estudio. Los pliegos fueron devueltos a PR y luego se distribuyeron duplicados a otras instituciones —v.gr., PRC.
M. clandestinum Phil., Anales Univ. Chile 85: 708 (1894). TIPO: [Chile, IX Región de La Araucanía, Cerros de Nahuelbuta:] Habitat in montibus Araucaniae Nahuelbuta dictis, I-1877, Philippi s.n. [lectotipo, designado por Calviño & al. (2016): SGO 053415 foto!; isolectotipo: SGO 041614 foto!) = Azorella spinosa (Ruiz & Pav.) Pers., Syn. Pl. 1: 303 (1805).
M. cryptanthum Clos in Gay, Fl. Chil. 3(1): 90-91 (1848-1849). TIPO: [Chile:] provincia de Coquimbo, cordilleras de Ovalle, Gay 515 (lectotipo, designado aquí: P 00115315 foto!; isolectotipos: P 00115314 foto!; P foto F neg. 28332!) = Azorella cryptantha (Clos) Reiche, Bot. Jahrb. Syst. 28(1): 17 (1899).
Se designa aquí el pliego recolectado por Gay 515, P 00115315, como lectotipo de M. cryptanthum entre sus duplicados, porque dicho pliego presenta mayor cantidad de material.
M. cuneatum Hook. & Arn., Bot. Beechey Voy.: 26 (1830). TIPO: [Chile:] Valparaíso, Cruckshanks 186 [lectotipo, designado por Calviño & al. (2016): K 000532301 foto!] = Azorella spinosa (Ruiz & Pav.) Pers., Syn. Pl. 1: 303 (1805).
M. dipterygia DC., Prodr. 4: 80 (1830). Asteriscium dipterygia (DC) Drude, Nat. Pflanzenfam. 3(8): 135 (1897). TIPO: Chile, Cordilleras, Haenke s.n. (lectotipo, designado aquí: PR 179033 foto!; isolectotipo: PRC 450112 foto!) = Gymnophyton polycephalum (Gillies & Hook.) Clos in Gay, Fl. Chil. 3(1): 103, lám. 32 fig. 1 (1848-1849). Se designa el pliego PR 179033 como lectotipo de M. dipterygia entre su duplicados, por ser el que presenta mayor abundancia de material. Véanse las observaciones hechas sobre etiquetas en M. angulatum.
M. integrifolium Hieron. Bol. Acad. Nac. Córdoba 4: 28-29 (1881). TIPO: [Argentina:] San Juan, Las Cuevas, I-1876, Echegaray s.n. (lectotipo, designado aquí: CORD 00003540!) = Azorella cryptantha (Clos) Reiche, Bot. Jahrb. Syst. 28(1): 16 (1899).
Se designa aquí el pliego recolectado por Echegaray s.n., CORD 00003540, como lectotipo de M. intregrifolium entre este y el sintipo de Hieronymus & Niederlein 309 —G 00359836, G foto F neg. 28333—, porque presenta mayor cantidad de material vegetativo y frutos.
M. isatidicarpum DC., Prodr. 4: 80 (1830). Asteriscium isatidicarpum (DC.) Hook & Arn., Bot. Misc. 3: 349 (1833). TIPO: [Chile:] Cordillera de Chile, Haenke s.n. (lectotipo, designado aquí: PR 179032 foto!; isolectotipo: PRC 450110 foto!) = Gymnophyton isatidicarpum (DC.) Mathias & Constance, Univ. Calif. Publ. Bot. 33(2): 145 (1962).
Se designa aquí el pliego PR 179032 como lectotipo de M. isatidicarpum dado que presenta mayor cantidad de material que su duplicado. Véanse las observaciones hechas sobre etiquetas en M. angulatum.
M. saniculaefolium (Lam.) Spreng., Umbellif. Prod.: 15 (1813). Hydrocotyle saniculaefolia Lam., Encycl. 3(1): 154 (1789). TIPO: [Uruguay:] Montevideo. Commerson s.n. (lectotipo, designado aquí: MPU 019658 foto!; isolectotipos: MPU 019656 foto!, MPU 019657 foto!, P00115473 foto!, P00115474 foto!, P00115475 foto!) = Diposis saniculaeolia (Lam) DC., Coll. Mém.: 38 (1829).
Se designa aquí el pliego recolectado por Commerson s.n., MPU 019658, como lectotipo de M. saniculaefolium, ya que es el pliego que presenta mayor cantidad de material. El pliego P 00115473 es equivalente en este sentido, pero en su etiqueta figura “Buenos Aires” en vez de Montevideo; lo hemos considerado isolectotipo, no obstante, ya que la especie no crece en Buenos Aires.
AGRADECIMIENTOSTOP
Los autores expresan su agradecimiento a los conservadores y al personal de los herbarios citados en el texto, por facilitar la consulta y el envío de ejemplares estudiados para este trabajo. A Ailén Santomé por las ilustraciones, a Susana Martínez por sus comentarios para mejorar versiones preliminares del manuscrito y a Enrique Rico y a un revisor anónimo, por sus importantes sugerencias. La primera autora agradece al CONICET, Argentina, por la concesión de la beca de doctorado necesaria para la realización de este proyecto. Este trabajo fue financiado por ANPCyT-FONCyT PICT 2011-1036, CONICET PIP 112-200901-00282 y la Universidad Nacional del Comahue PIN B180 de Argentina a CC y CE.
REFERENCIAS BIBLIOGRÁFICASTOP
| ○ |
Aubert, S., Boucher, F., Lavergne, S., Renaud, J. & Choler, P. 2014. 1914-2014: A revised worldwide catalogue of cushion plants 100 years after Hauri and Schröter. Alpine Botany 124: 59-71. https://doi.org/10.1007/s00035-014-0127-x. |
| ○ |
Badano, E. 2006. Asociaciones de especies a plantas en cojín: sus consecuencias sobre la diversidad de especies vegetales en comunidades alto-Andinas. Ecosistemas 15(1): 109-112. |
| ○ |
Baillon, H.E. 1880. Ombelliferes. In: Baillon, H.E. (ed.), Histoire naturelle des plantes 7: 84-256. Hachette, París. |
| ○ |
Barboza, G.E., Cantero, J.J., Nuñez, C., Pacciaroni, A. & Ariza Espinar, L. 2009. Medicinal plants: a general review and a phytochemical and ethnopharmacological screening of the native Argentine Flora. Kurtziana 34(1-2): 7-365. |
| ○ |
Beentje, H. 2010. The Kew Plant Glossary and illustrated dictionary of plants terms. Royal Botanic Gardens, Kew. |
| ○ |
Bentham, G. 1867. Umbelliferae. In: Bentham, G. & Hooker, J.D. (eds.), Genera Plantarum 1: 859-931. Reeve, Londres. |
| ○ |
Brummitt, R.K. & Powell, C.E. 1992. Authors of plant names: a list of authors of scientific names of plants, with recommended standard forms of their names, including abbreviations. Royal Botanic Gardens, Kew. |
| ○ |
Cabrera, A.L. 1971. Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica 14(1-2): 1-50. |
| ○ |
Cabrera, A.L. & Willink, A. 1980. Biogeografía de América Latina. Secretaria General de la Organización de los Estados Americano, Washington DC. |
| ○ |
Calviño, C.I., Fernández, M. & Martínez, S. 2016. Las especies del género Azorella (Azorelloideae, Apiaceae) con distribución extra-Argentina. Darwiniana 4(1): 57-82. https://doi.org/10.14522/darwiniana.2016.41.681. |
| ○ |
Cavagnaro, F.P., Golluscio, R.A., Wassner, D.F. & Ravetta, D.A. 2003. Caracterización química de arbustos patagónicos con diferente preferencia por parte de los herbívoros. Ecología Austral 13: 215-222. |
| ○ |
Cavanilles, A.J. 1799. Selinum. In: Cavanilles, A.J. (ed.), Icones et descriptions plantarum, qua aut spont in Hispania crescunt, aut in hortis hospitantur 5: 58-60. Biblioteca Valenciana, Madrid. |
| ○ |
Clos, D. 1848-1849. Umbelíferas. In: Gay, C. (ed.), Historia física y política de Chile, Botánica 3: 61-145. Museo de Historia Natural de Santiago, Santiago de Chile. |
| ○ |
Constance, L. 1965. Some subtractions from the Umbelliferae of South America. Annals of the Missouri Botanical Garden 52(3): 274-280. https://doi.org/10.2307/2394793. |
| ○ |
Constance, L., Chuang, T.I. & Bell, C.R. 1971. Chromosome numbers in Umbelliferae IV. American Journal of Botany 58(6): 577-587. https://doi.org/10.2307/2441041. |
| ○ |
Constance, L., Chuang, T.I. & Bell, C.R. 1976. Chromosome numbers in Umbelliferae V. American Journal of Botany 63(5): 608-625. https://doi.org/10.2307/2441824. |
| ○ |
Constance, L. 1988. Umbelliferae. In: Correa, M.N. (ed.), Flora Patagónica 8(5): 310-379. Colección Científica del Instituto Nacional de Tecnología Agropecuaria (INTA), Buenos Aires. |
| ○ |
Damascos, M.A., Barthélémy, D., Ezcurra, C., Martínez, P. & Brion, C. 2008. Plant phenology, shoot growth, and branching pattern in Mulinum spinosum (Apiaceae), a cushion shrub of the arid Patagonian steppe of Argentina. Journal of Arid Environments 72: 1977-1988. https://doi.org/10.1016/j.jaridenv.2008.07.001. |
| ○ |
De Candolle, A.P. 1830. Umbelliferae. In: De Candolle, A.P. (ed.), Prodromus systematis naturalis regni vegetabilis 4: 55-250. Treüttel & Würtz, París. |
| ○ |
De Mösbach, E.W. 1992. Botánica indígena de Chile. Museo Chileno de Arte Precolombino. Fundación Andes, Chile. |
| ○ |
Drude, C.G.O. 1897-1898. Umbelliferae. In: Engler, A. & Prantl, K. (eds.), Die naturlichen Pflanzenfamilien 3(8): 63-250. Wilhelm Engelmann, Lipsia. |
| ○ |
Echegaray, D.S. 1881. Umbelíferas. In: Hieronymus, G. (ed.), Boletín de la Academia Nacional de Ciencias en Córdoba 4: 26-29. Guillermo Kraft, Córdoba. |
| ○ |
Estomba, D., Ladio, A. & Lozada, M. 2005. Plantas medicinales utilizadas por una comunidad mapuche en las cercanías de Junín de los Andes, Neuquén. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 4: 107-112. |
| ○ |
Faggi, A.M. 1998. Listado preliminar de especies del Parque Nacional Los Alerces. In: Administración de Parques Nacionales. Sistema de Información de Biodiversidad, Argentina [http://www.sib.gov.ar]. |
| ○ |
Fernández, M., Ezcurra, C. & Calviño, C.I. 2014. Mulinum ulicinum (Azorelloideae, Apiaceae), nueva cita para Chile. Gayana Botánica 71(2): 278-280. https://doi.org/10.4067/S0717-66432014000200009. |
| ○ |
Fernández, M., Martínez, S. & Calviño, C.I. 2016a. Proposal to conserve the name Selinum microphyllum (Mulinum microphyllum) (Apiaceae: Azorelloideae) with a conserved type. Taxon 65(2): 396-397. https://doi.org/10.12705/652.28. |
| ○ |
Fernández, M., Ezcurra, C. & Calviño, C.I. 2016b. Morphology, fruit anatomy and taxonomy of the South Andean Genus Laretia (Azorelloideae, Apiaceae). Systematic Botany 41(3): 807-812. https://doi.org/10.1600/036364416X692370. |
| ○ |
Fernández, M., Ezcurra, C. & Calviño, C.I. 2017a (in press). Species limits and morphometric and environmental variation within the South Andean and Patagonian Mulinum spinosum species-group (Apiaceae-Azorelloideae). Systematics and Biodiversity 15. https://doi/full/10.1080/14772000.2016.1273975. |
| ○ |
Fernández, M., Ezcurra, C. & Calviño, C.I. 2017b. Chloroplast and ITS phylogenies to understand the evolutionary history of southern South American Azorella, Laretia and Mulinum (Azorelloideae, Apiaceae). Molecular Phylogenetics and Evolution 108: 1-21. |
| ○ |
Ferreyra, M., Ezcurra, C. & Clayton, S. 2006. Flores de Alta Montaña de los Andes Patagónicos. LOLA, Buenos Aires. |
| ○ |
Gillies, J. & Hooker, W.J. 1830. On the plants of the natural order Umbelliferae. In: Hooker, W.J. (ed.), Botanical Miscellany 1: 323-335. John Murray, Londres. |
| ○ |
Grisebach, A.H.R. 1874. Umbelliferae. In: Grisebach, A.H.R (ed.), Abhandlungen der Königlichen Gesellschaft der Wissenschaften zu Göttingen 19: 153-156. Der dieterichschen buchhandlung, Gotinga. |
| ○ |
Hijmans R.J., Guarino, L., Bussink, C., Mathur, P., Cruz, M., Barrantes, I. & Rojas, E. 2009. DIVA-GIS 7.5. A geographic information system for the analysis of species distribution data [http://www.diva-gis.org]. |
| ○ |
Hiroe, M. 1979. Mulinum. In: Hiroe, M. (ed.), Umbelliferae of World: 1728-1732. Botanical Institute, Japón. |
| ○ |
Hunziker, A.T. 1960. Catálogo de los tipos “grisebachianos” conservados en Córdoba. Boletín de la Academia Nacional de Ciencias en Córdoba 41: 283-421. |
| ○ |
Johnston, I.M. 1928. The botanical activities of Thomas Bridges. Contributions from the Gray Herbarium of Harvard University 81: 98-106. |
| ○ |
Kuntze, C.E.O. 1891-1893. Umbelliferae. In: Kuntze, C.E.O. (ed.), Revisio generum plantarum 3(2): 110-115. A. Felix, Lipsia. |
| ○ |
Macloskie, G. 1905. Umbelliferae. In: Scott, W.B. (ed.), Reports of the Princeton University expeditions to Patagonia 1869-1899 8(2): 619-643. Schweizerbart, Estucardia. |
| ○ |
Martínez, S. 1989. El género Azorella (Apiaceae-Hydrocoyloideae) en la Argentina. Darwiniana 29(1-4): 139-178. |
| ○ |
Martínez, S. 2003. Umbelliferae. In: Kiesling, R. (ed.), Flora de San Juan 2: 214-234. Estudio Sigma, Buenos Aires. |
| ○ |
Martínez, S. 2008. Apiaceae. En: Zuloaga, F.O., Morrone, O. & Belgrano, M.J. (eds.), Catálogo de las Plantas Vasculares del Cono Sur 2: 1056-1090. Missouri Botanical Garden Press, San Luis. |
| ○ |
McNeill, J., Barrie, F.R., Buck, W.R., Demoulin, V., Greuter, W., Hawksworth, D.L., Herendeen, P.S., Kanpp, S., Marhhold, K., Prado, J., Prud’homme van Reine, W.F., Smith, G.F., Wiersema, J.H. & Turland, N.J. 2012. International Code of Nomenclature for algae, fungi and plants (Melbourne Code). Regnum Vegetabile 154. Koeltz Scientific Books, Königstein. |
| ○ |
Moreno, F.P. 1876-1877. Viaje a la Patagonia Austral. Ediciones Solar, Buenos Aires. |
| ○ |
Moreno, F.P. 1897. Apuntes preliminares sobre una excursión a los territorios del Neuquén, Río Negro, Chubut y Santa Cruz. Talleres de publicaciones del Museo de La Plata, La Plata. |
| ○ |
Munizaga, C. & Gunckel, H. 1958. Notas etnobotánicas del pueblo atacameño de Socaire. Centro de estudios antropológicos. Universidad de Chile, Chile. |
| ○ |
Muñoz, O., Montes, M. & Wilkomirsky, T. 1999. Plantas medicinales de uso en Chile. Química y Farmacología. Editorial Universitaria, Chile. |
| ○ |
Nicolas, A.N. & Plunkett, G.M. 2009. The demise of subfamily Hydrocotyloideae (Apiaceae) and the re-alignment of its genera across the entire order Apiales. Molecular Phylogenetics and Evolution 53: 134-151. https://doi.org/10.1016/j.ympev.2009.06.010. |
| ○ |
Nicolas, A.N. & Plunkett, G.M. 2012. Untangling generic limits in Azorella, Laretia, and Mulinum (Apiaceae: Azorelloideae): Insights from phylogenetics and biogeography. Taxon 61(4): 826-840. |
| ○ |
Niemeyer, H.M. 1995. Biologically Active Compounds from Chilean Medicinal Plants. In: Arnason, J.T., Matta, R. & Romeo, J.T. (eds.), Phytochemistry of Medicinal Plants 29: 137-159. Springer, México. https://doi.org/10.1007/978-1-4899-1778-2_7. |
| ○ |
Nuñez, C., Aizen, M. & Ezcurra, C. 1999. Species associations and nurse plant effects in patches of high-Andean vegetation. Journal of Vegetation Science 10: 357-364. https://doi.org/10.2307/3237064. |
| ○ |
Persoon, C.H. 1805. Mulinum. In: Persoon, C.H. (ed.), Synopsis Plantarum 2: 309. Parisiis Lutetiorum, París. |
| ○ |
Philippi, R.A. 1860. Umbelliferae. In: Philippi, R.A. (ed.), Viaje al desierto de Atacama 198-200. Eduardo Anton, Santiago de Chile. |
| ○ |
Philippi, R.A. 1864. Mulinum. In: Schlechtendal, D.F.L (ed.), Linnaea 33: 90. F. Dümmler, Berlín. |
| ○ |
Philippi, R.A. 1893-1894. Umbelliferae. In: Philippi, R.A. (ed.), Anales de la Universidad de Chile, Memorias Científicas y Literarias 85: 707-710. Imprenta Cervantes, Santiago de Chile. |
| ○ |
Pontiroli, A. 1965. Mulinum. In: Cabrera, A.L. (ed.), Flora de la Provincia de Buenos Aires 4: 387. Colección Científica del INTA, Buenos Aires. |
| ○ |
Rauh, W. 1939. Über polsterförmigen Wuchs, ein Beitrag zur Kenntnis der Wuchsformen der höheren Pflanzen. Nova Acta Leopoldiana 7: 268-508. |
| ○ |
Ravenna, P. 2008. Studies in Apiaceae I, a new Mulinum species, notes on the genuine Mulinum spinosum (Cav.) Pers., and revalidation of M. proliferum Phil. and M. ovalleanum Phil. Onira 11: 56-58. |
| ○ |
Reiche, K. 1899. Umbelíferas. In Reiche, K. (ed.), Anales de la Universidad de Chile 104: 767-842. Editorial Universitaria, Santiago de Chile. |
| ○ |
Seoane, N.F., Ochoa, J., Borrelli, L., Severino, M.E., Taddeo, H., Giraudo, C. & Andrade Gamboa, J. 2011. Mulinum spinosum y la carne de cordero: detección de su presencia en ovinos vivos. Archivos de Zootecnia 60: 283-292. https://doi.org/10.4321/S0004-05922011000200013. |
| ○ |
Skottsberg, C.J.F. 1916. Mulinum. In: Skottsberg, C.J.F. (ed.), Kungliga Svenska Vetenskapsakademiens Handlingar 56(5): 278-280. L.L. Grefing, Estocolmo. |
| ○ |
Spegazzini, C.L. 1899. Nova addenda ad Floram Patagonicam, Anales de la Sociedad Científica Argentina 48: 44-59. |
| ○ |
Systematics Association Committee for the Descriptive Biological Terminology. 1962. II. Terminology of simple symmetrical plane shapes (Chart 1). Taxon 11: 145-156. https://doi.org/10.2307/1216718. |
| ○ |
Thiers, B. 2015. Index Herbariorum: a global directory of public herbaria and associated staff. [http://sweetgum.nybg.org/ih/]. |
| ○ |
Tseng, C.C. 1967. Anatomical studies of flower and fruit in the Hydrocotyloideae (Umbelliferae). University of California Publications in Botany 42: 1-59. |
| ○ |
Villagrán, C., Castro, V., Sanchez, G., Romo, M., Latorre, C. & Hinojosa, L.F. 1998a. La tradición surandina del desierto: Etnobotánica del área del Salar de Atacama (Provincia de El Loa, Región de Antofagasta, Chile). Estudios atacameños 16: 7-105. |
| ○ |
Villagrán, C., Castro, V. & Sanchez, G., 1998b. Etnobotánica y percepción del paisaje en Caspana (Provincia de El Loa, Región de Antofagasta, Chile): ¿Una cuña atacameña en el Loa Superior? Estudios atacameños 16: 107-170. |
| ○ |
Webb, C.J. 1981. Andromonoecism, protandry, and sexual selection in Umbelliferae. New Zealand Journal of Botany 19: 335-338. https://doi.org/10.1080/0028825X.1981.10426389. |
| ○ |
Weddell, H.A. 1857. Umbelliferae. In: Weddell, H.A. (ed.), Chloris Andina. Essai d’ une flore de la région alpine des Cordillères de l ’Amérique du Sud 2: 186-206. Bertrand, París. |
| ○ |
Wolff, H. 1921. Mulinum. Repertorium Specierum Novarum Regni Vegetabilis 17: 441-442. https://doi.org/10.1002/fedr.19210171931. |
| ○ |
Zech, J.C. 1992. Systematics of the genus Mulinum Pers. (Apiaceae, Hydrocotyloideae, Mulineae). PhD dissertation, The Ohio State University, Ohio. |
| ○ |
Zech, J.C. 1995. Nomenclatural novelties on Mulinum (Apiaceae). Kurtziana 24: 192. |
APÉNDICESTOP
APÉNDICE 1. Índice de recolectores y número de recolector. En el caso de no poseer número se indica el número del herbario en el que se encuentra depositado o, en su defecto, solo el herbario. Entre paréntesis se indica el número de orden del taxon en el tratamiento taxonómico.
Sine col. SI 165406 (7); SI 165407 (7); SI 165409 (7); SI 165410 (7); Aedo 6997 (2); Ambrosetti 1220 (3); 1243 (3); Ameghino 25 (4); LP 002796 (10); Ancibor 119 (10); Ancibor & Mujica 32 (9); Ancibor & al. 90214 (1); Arancio 92-640 (2); 10647 (2); Arenas 947 (8); Arroyo & al. 20 (7); 147 (4); 210 (4); 223 (10); 274 (4); 358 (10); 382 (10); 421 (4); 426 (9); 462 (7); 508 (10); 562 (10); 2252 (10); 2384 (10); 2610 (10); 202079 (7); 841002 (10); 870276 (10); 94140 (2); 94010 (2); 97268 (9); 97290 (9); 97385 (9); Arnow 3780 (6); Ayarda 363 (9); Ayarda & Muruaga 173 (9); Ayestaran 10008 (6); Bach 304 (7); Baeza 268 (7); Baeza & al. 2981 (5); Barboza & al. 1207 (7); 2550 (8); 2555 (8); 2576 (8); 2897 (8); 2925 (8); 3163 (8); 3350 (8); Barboza & Cantero 3925 (8); 3932 (8); Blake 457 (5); Beck 14141 (9); 21726 (8); Beetle & Soriano 509 (10); Belgrano 69 (7); Berg CORD (4); Bernardello & Figueroa 312 (7); 313 (7); Bernardello & Moscone (5); Biganzoli & al. 2287 (4); 2380 (4); Biurrun 5757 (3); Boelcke & Correa 6985 (5); 6987 (5); 6988 (5); 6989 (5); 6990 (5); 7056 (5); Boelcke & Hunziker 3484 (6); 3512 (6); 3572 (6); Boelcke & al. 1728 (6); 1760 (6); 4217 (1); 4301 (7); 4624 (1); 7574 (6); 8818 (7); 9940 (3); 10033 (1); 10064 (1); 10205 (7); 10236 (7); 10309 (7); 10648 (7); 10970 (7); 11228 (1); 11378 (5); 11610 (1); 11613 (5); 11500 (5); 11633 (1); 12229 (4); 12281 (4); 12760 (4); 12801 (4); 12912 (10); 13564 (7); 13761 (7); 13897 (5); 13972 (7), 13974 (7); 14121 (5); 14179 (5); 14246 (7); 14298 (5); 15368 (4); 15538 (7); 15747 (7); 15927 (1); Bonifacino & Donato 733 (4); Bonifacino & al. 2971 (4); 2982 (10); 2983 (7); Borgnia 32 (8); Bottino 247 (7); Bravo LIL 539722 (9); Bridges 1842 (1); Buchtien SI 10263 (7); Burkart 5344 (9); 6221 (7); 6642 (6); Burkart & Múlgura 28191 (7); Burkart & Troncoso 11811 (9); Burmeister BAB 6059 (4); 6061 (4); Cabrera 34 (7); 35 (7); 4283 (6); 4819 (7); 5507 (7); 5507 (7); 5767 (7); 6177 (7); 6720 (7); 8353 (9); 8378 (9); 8672 (9); 8790 (1); 9069 (9); 9214 (9); 9221 (9); 9431 (8); 11067 (7); 11132 (7); 11210 (6); 18713 (7); 25964 (5); 27458 (8); 31775 (9); 32468 (8); 32836 (7); 32874 (5); 32965 (7); 33021 (6); 33177 (4); 33270 (7); 33279 (7); 33331 (7); Cabrera & Crisci 19112 (7); 19191 (5); Cabrera & Job 39 (6); Cabrera & al. 17734 (8); 18947 (8); 18964 (9); 22540 (9); 27181 (8); 33116 (6); 33128 (6); 33250 (10); Calviño & al. 106 (7); 700 (7); 701 (6); 702 (7); 704 (6); 706 (5); 709 (7); 721 (5); 726 (7); 727 (5); 734 (7); 745 (7); 746 (5); 747 (6), 748 (6); 751 (5); 752 (7); 753 (5); 757 (3); 788 (2); 790 (2); 792 (9); 793 (9); 794 (9); 797 (9); 798 (9); 808 (8); 809 (8); 813 (8); 814 (8); 816 (8); 817 (8); 817bis (8); 818 (8); 819 (8); Canessa 53 (7); Carette LP 23378 (4); 290 (1); 2515 (1); Castellanos BA 28/291 (8); BA 28/292 (8); BA 6111 (4); BA 6117 (10); BA 6119 (10); LIL 15501 (9); BA 36808 (1); BA 27/846 (9); LIL 420516 (9); Castillón 7188 (9); Chandler & Bayer 1105 (1); 1111 (7); 1114 (1); Charpin & Ahumada 20673 (8); Charpin & Lazare 24022 (8); 24050 (8); Chiapella 1667 (7); Chiapella & al. 122 (7); 144 (7); 192 (7); 223 (5); 284 (1); 444 (7); 528 (7); 544 (1); Chiapella & Clayton 13 (7); 625 (7); Chiapella & Puntieri 13 (7); Cittadini 52bis (4); Claren 11702 (8); Cocucci 58 (3); 4124 (7); 4125 (7); 4127 (7); 4128 (7); 4131 (7); 4132 (7); 4133 (7), 4134 (7); Cocucci & Sérsic 2362 (7); 2466 (10); 2482 (7); 4120 bis (7); 4121 (7); Cocucci & al. 2211 (7), 2112 (7); Constance 3507 (7); 3506 (7); 3516 (7); 3856 (9); Constance & Cabrera 3854 (8); Constance & Sparre 3577 (7); 3579 (7); Constance & al. 3544 (7); Cordi & al. BCRU (6); Cordi & al. BCRU (7); Correa & Nicora 3335 (4); Correa & al. 2586 (4); 2598 (4); 2651 (4); 3817 (6); 3836 (4); 3936 (7); 3970 (6); 3972 (7); 3985 (4); 3997 (6); 4011 (4); 4063 (4); 4065 (7); 4085 (6); 4090 (10); 4096 (7); 4136 (7); 4152 (6); 4181 (7); 4202 (7); 5815 (5); 5817 (5); 5853 (5); 5855 (5); 5912 (5); Cottet 8 (6); Covas 10827 (1); 10828 (1); 10913 (1); Cuezzo & Barkley 20Mz383 (1); Cumming 1188 (1); Daniels 22 (6); Dawson 1032 (7); 1271 (6); De la Sota 2725 (9); Del Vitto & al. 36530 (7); Descole 2420 (7); Diem 1108 (5); 3349 (5); 1841 (5), 1844 (5); Dinelli 542 (8); Domínguez 286 (10); Donat 60 (4); 61 (4); Donoso & Schwabe 2633 (5); Dusén 5304 (4); Elechosa & al. BAB (7); Eskuche 299-1 (5); 599-19 (5); 927b (5); 926 (6); 928 (5); 929 (5); 1448 (5); Eyerdan & al. 23963 (4); Ezcurra 786 (5); 956 (6); 3115 (5); 3502 (7); Ezcurra & al. 703 (6); 727 (7); 799 (5); 1051 (5); 1259 (7); 2433 (6); 2482 (5); 2581 (1); 2656 (5); 2666 (1); 2909 (5); 2974 (5); 2980 (5); 3000 (5); 3291 (7); Ezcurra & Bianchi 1908 (6); Ezcurra & Galende 977 (6); 1008 (7); Ezcurra & Quiroga 3163 (5); Fabris & Crisci 7595 (7); Fabris & Schwabe 4918 (7); Faggi BA 78048 (6); BA 78052 (6); Faggi & Lopéz 224 (10); Fasanella 8340 (7); Fernández 6 (7); 7 (7); 12 (4); 13 (4); 14 (7); 17 (7); 20 (4); 22 (4); 24 (7); 25 (7); 27 (4); 30 (10); 32 (4); 34 (4); 40 (4); 41 (4); 44 (7); 50 (8); 51 (8); 57 (8); 60 (8); Ferrando 30 (7); Ferreyra 216 (5); 330 (5); 342 (5); 390 (5); 412 (6); Ferreyra & Chiapella 578 (5); Ferreyra & Clayton 198 (7); 291 (7); Ferreyra & Ezcurra 610 (7); Ferreyra & Siracussa 622 (5); Ferruglio 30/1873 (4); 30/1878 (4); 34744 (4); 34745 (4); 34747 (4); 34748 (4); Fiebrig 2601 (8); Forcone & al. 10000 (4); 10001 (4); 10002 (4); 10003 (4); 10004 (4); 10005 (4); 10006 (4); 10007 (4); 10009 (6); Fortunato 5587 (7); Fries 840 (9); Funes & Arenas 22 (7); Garaventa 4361 (7); Garcés 25 (6); Geisse SGO 000003751 (7); SGO 000003753 (7); Gerling LP 023376 (8); Giacanelli LIL 64434 (9); Gillies E 00000042 (1); E 00000043 (1); Gómez 205a (7); Gonzalez 22 (8); 210 (7); Grau LIL 519837 (8); Greer 1 (5); Grondona 2202 (10); Grossfeld BCRU (7); Guerrido & Fernández 831 (10); Gutierrez & Lopez 46 (2); Haene 1625 (3); Haenke HAL 26839 (7); Halloy 39 (10); 583 (8); Hicken 6081 (7); 116323 (7); Hicken & Hauman 138 (4); 253 (4); 340 (4); 360 (4); 438 (4); 445 (4); Hieronymus & Niederlein 360 (8); 792 (8); 819 (8); Hollermayer 489 (5); 1724 (5); Hosseus 1151 (8); Huek 511 (2); Hunziker 4145 (9); 1884 (8); 2099 (8); 6993 (5); 10605 (9); 22843 (8); Hunziker & al. 11310 (1); Illin 8 (4); LP 10612 (6) BAB 6043 (4); BAB 6045 (4); 257 (6); 6038 (4); 6058 (6); Jörgensen 1692 (8); Kiesling 4532 (3); 6763 (1); 7688 (9); 10408 (9); 10412 (3); 3228 (9); Kiesling & al. 8068 (7); Koslowsky 94 (4); 95 (7); 145 (4); 1314 (10); 1318 (4); 1319 (4); 12353 (10); 12371 (4); Kragkievich 3949 (4); Krapovickas 3212 (9); 6184 (8); 6250 (8); 6337 (8); 10655 (8); Künhnemann 471 (6); 552 (6); Kuntze NY 00073906 (7); Kurtz CORD (3); 3387 (7); 5622 (7); 5756 (1); 5904 (7); 5905a (1); 7505 (7); 7507 (7); 9372 (7); 9463 (3); 9652 (3); 9816 (3); 13655 (8); Lahitte 52011 (6); Landero 643 (10); 699 (10); Landrum & Landrum 8410 (7); Latorre & al. 206 (2); Latour 205b (7); Lechler 3000 (6); Lechler P 03254892 (6); Leuenberger 4060 (7); 4118 (10); Llera & Torres 1868-7 (4); 1921-11 (4); Lorentz 136 (8); Lorentz K 000529780 (8); Lorentz SI 000739 (8); Losert BCRU (4); BCRU (4); 11 (4); Lotti de Stange 133 (7); 139 (7); Luján 46 (3); 64 (3); Luti & al. 5743 (7); Magens 918 (10); Marticorena & Matthei 629 (7); 761 (7); 937 (7); Marticorena & al. 1398 (5); Martínez Carretero 144 (7); 1122 (9); Méndez 9888 (7); 10462 (7); 10473 (7); Méndez & Willoud 31495 (1), 31516 (1); 32975 (7); Miche 171/80/28.3 (5); Mieres CONC 166141 (1); Mihoc & al. 4590 (7); 4600 (5); 6921 (7); Milner 22426 (7); Mintzer 60101 (9); 60102 (9); Montaña 532 (2); Montenegro BCRU (4); Montero 7080 (7); 8082 (7); 11308 (7); Montiel LIL 246699 (5); Moreira & Muñoz 300 (9); 318 (9); Morrison 16915 (7); Morrone 5702 (7); 6135 (6); Naranjo & al. 906 (7); Née CONC 29919 (7); F 68085 (7); MA 310354 (7); Nicora 3668 (7); Norman 3126 (9); Novara 6347 (8); 13400 (9); Novara & Charpin 10434 (8); Novara & Neumann 9775 (8); Novara & Osten 10078 (8); Nuñez & Amoedo 185 (6); Nuñez & Daniels 156 (5); Nuñez & Ezcurra 84 (7); Nuñez 47 (5); 751 (9); 763 (9); O´Donell 3560 (4); 3663 (4); 3799 (4); 3936 (4); Onell 2049 (5); Otto BAB 8003 (5); Pastore 68 (7); Patiño 7917 (1); Pedersen 14469 (7); Pedreros 50 (2); Pérez 2 (2); Perez Moreau BA 30/147 (9); 18663 (6); 23311(7); 23314 (7); 45363 (6); 45379 (5); 45386 (5); 48694 (6); 48695 (6); 48689 (6); 49140 (5); 49142 (6); 49902 (5); 49907 (5); 50371 (5); Pestalozzi 474 (8); Perrone BA 30254 (1); Pfister 946 (7); 7132 (7); 9374 (2); CONC 18487 (7); Pfister & Ricardi 417 (7); Philippi K 000529783 (7); K 000529782 (7); K 000529786 (2), K 000529787 (2); BM 001042508 (7); F 0BN003451 (7); Plotrick 49 (7); Puntieri 57 (6); 257 (7); 336 (6); BCRU (7); BCRU (6); Puntieri & Chiapella 85 (6); Raffo 139 (4); Rahmer SGO 41607 (5); Ricardi 3164 (7); 3168 (7); Ricardi & Marticorena 5666 (5); Ricardi & Matthei 282 (4); Ricardi & al. 758 (7); Rodriguez 32 (9); 293 (8); 1338 (9); 3210 (2); 3232 (2); Roig 3029 (7); 3186 (3); 4232 (7); 4855 (7); 5274 (9); 11117 (3); MERL 45581 (3); 11949 (9); Roig & Méndez 9887 (4); 15096 (6); Roig & Ruiz Leal 3319 (3); 5234 (9); 20166 (3); 45571 (1); Roig & al. 46839 (7); Roivainen SGO 79316 (6); Rossi 317 (1); Rosso BCRU (6); Rossow & al. 1876 (5); 2374 (7); Rubulis 265a (5); Ruiz Leal & Roig 15654 (1); 18833 (9); 20719 (3); 23635 (9); 23676 (9); Ruiz Leal & al. 1855 (9); 1983 (1); 2021 (7); 2034 (9); 2887 (7); 3155 (1); 3169 (1); 3184 (9); 3619 (1); 7182 (9); 7183 (1); 7828 (1), 11682 (1); 14578 (1); 14784 (10); 16082 (4); 15763 (1); 17563 (1), 20244 (6); 21045 (9); 21439 (1); 21988 (7); 22387 (7); 23156 (7); 23719 (7); 24082 (4); 24109 (4); 24172 bis (7); 24218 (4); 24225 (10); 24357 (7); 24437 (7); 24532 (7); 24544 (7); 25591 (6); 25663 (7); 25807 (5); 26423 (10); 26425 (4); 26434 (10); 26451 (10); 26772 (6); 26905 (4); 27020 (10); Ruthsatz 7 (9); 35 (8); 40 (9); 41 (9); 44 (9); 47 (9); 57 (9); 126 (9); 135 (9); 136 (9); 166 (8); 204 (9); 210 (9); 225 (9); 241 (8); 275 (9); 336 (9); 363 (8); 507 (8); 9073 (9); 9199 (8); Sage 1703210 (7); 1765128 (7); Salazar 15209 (8); Salinas 2180 (1); Schickendantz 334 (8); Schlegel 2476 (7); 5124 (2); Schwabe & al. 1037 (9); Scott de Birabén 25 (7); 187 (7); 450 (7); 518 (7); Semper MERL 45540 (9); 10091 (3); 11855 (3); 13959 (9); Sermolli & Bizzarri 7372 (7); sin col. LIL 64433 (9); LP 22537 (4); Silvestre 5588 (4); 5987 (4); Sleumer 1178 (10); 1358 (10); 1460 (5); 1874 (9); 3311 (9); 3365 (9); Skottsberg 855 (4); Sleumer & Vervoorst 2578 (8); Soriano 231 (5); 1320 (6); 1986 (6); 2025 (4); 2160 (10); 2193 (10); 2257 (4); 2426 (4); 2508 (6); 3072 (10); 3788 (6); 4795 (10); 5076 (4); 5098 (4); 5099 (4); Sosa 11 (5); 27132 (1); Spegazzini ex LPS 784 LP (6); SI 6911 (7); Stecconi BCRU (6); Stuessy & Ruiz 12775a (7); Stuckert 2834 (7); 15125 (8); 17894 (9); 18118 (6); 23459 (7); Stuessy & al. 6892 (10); 10129 (5); 10333 (1); 10162 (5); Suárez 821 (4); Tammone 27 (6); Teague LIL 191353 (6); Teillier 3028 (2); 4556 (1); Tessleff 5304 (4); Troiani & Steibel 15470 (6); 15490 (6); 15715 (6); Troncoso 9310 (7); Urtubey & Baztarrica 579 (7); 634 (7); Varela & Del Castillo 1362 (9); Venturi 10279 (8); 80532 (4); Villagrán 8757 (2); Villagrán & Arroyo CONC 55941 (7); CONC 55947 (7); Villagrán & al. 7912 (5); 8128 (1); Villamil & al. 2228 (10); 2795 (5); von Thüngen 14 (4); 97 (4); 103 (4); Weigend & al. 6864 (5); Werdermann 268 (7); 613 (1); 1009 (2); West 4757 (6); Wilczek LAU (1); Zavala-Gallo & al. 213 (4); Zech & Contogiogakis 2 (7); 6 (7); 7 (7); 8 (7); 21 (4); 24 (10); Zöllner 2293 (5); 5208 (5); 7587 (5); Zuloaga 11928 (8).
APÉNDICE 2. Gráficos de caja para los caracteres cuantitativos utilizados en la clave dicotómica. Caracteres vegetativos: se representan la mediana, los percentiles del 25% y 75%, y las barras de error con los percentiles del 10% y 90%; los círculos sin relleno corresponden a valores atípicos u outliers.
APÉNDICE 2. (continuación). Gráficos de caja para los caracteres cuantitativos utilizados en la clave dicotómica. Caracteres reproductivos: Se representan la mediana, los percentiles del 25% y el 75% y las barras de error con los percentiles del 10% y el 90%; los círculos sin relleno corresponden a valores atípicos o outliers.