Cytogenetic relationships within the Maghrebian clade of Festuca subgen. Schedonorus (Poaceae), using flow cytometry and FISH
David Ezquerro-López1, David Kopecký2 & Luis Á. Inda1*
1Grupo BIOFLORA, Escuela Politécnica Superior de Huesca, Instituto Agroalimentario de Aragón–IA2, Universidad de Zaragoza, 22071 Huesca, Spain; lainda@unizar.es
2Centre of the region Haná for Biotechnological and Agricultural Research, Institute of Experimental Botany, Šlechtitelů 31, Olomouc-Holice, 78371, Czech Republic
* Corresponding author
| |
Abstract
Ezquerro-López, D., Kopecký, D. & Inda, L.Á. 2017. Cytogenetic relationships within the Maghrebian clade of Festuca subgen. Schedonorus (Poaceae), using flow cytometry and FISH. Anales Jard. Bot. Madrid 74(1): e052.
Festuca subgen. Schedonorus is a group of broad-leaved fescues, which can be divided into two clades: European and Maghrebian. We employed fluorescent
in situ hybridization —FISH— with probes specific for 5S and 35S ribosomal DNA and genome size estimation using flow cytometry to
shed light on the determination of possible parental genomes of polyploid species of the Maghrebian clade. Our results indicate
that octoploid F. arundinacea subsp. atlantigena probably originated from crossing of the tetraploids F. arundinacea subsp. fenas —2n = 4x = 28— and F. mairei —2n = 4x = 28— followed by whole genome duplication. However, a large reconstruction of karyotype and genome downsizing has been revealed.
Similarly, hexaploid F. arundinacea subsp. corsica presumably resulted from the interspecific hybridization of the diploid F. pratensis and tetraploid F. arundinacea subsp. fenas. Several scenarios on the origin of decaploid F. arundinacea var. letourneuxiana are discussed. This study contributed to our knowledge on the phylogeny of broad-leaved fescues and provided new information
on the karyotypes —chromosome numbers, ploidy levels and numbers and positions of rDNA loci— using FISH and genome size estimations
using flow cytometry in selected taxa of this important grass genus.
Keywords: Broad-leaved fescues,
Festuca subgen. Schedonorus,
FISH,
genome size,
rDNA.
|
| |
Resumen
Ezquerro-López, D., Kopecký, D. & Inda, L.Á. 2017. Relaciones citogenéticas en el clado magrebí de Festuca subgen. Schedonorus (Poaceae), mediante la utilización de citometría de flujo y FISH. Anales Jard. Bot. Madrid 74(1): e052.
Festuca subgen. Schedonorus es un grupo de festucas de hojas anchas que se divide tradicionalmente en dos clados, uno europeo y otro magrebí. Mediante
hibridación in situ fluorescente —FISH— con sondas específicas para las regiones ribosómicas 5S y 35S en su cariotipo y estimaciones de tamaño
genómico mediante citometría de flujo se intentó determinar los posibles genomas parentales de las especies poliploides del
clado magrebí. Nuestros datos indican que la especie octoploide F. arundinacea subsp. atlantigena probablemente se originó a partir del cruce de los tetraploides F. arundinacea subsp. fenas —2n = 4x = 28— y F. mairei —2n = 4x = 28— seguido de la duplicación del genoma. Sin embargo, también se ha detectado una reconstrucción del cariotipo y una reducción
del tamaño genómico. De forma similar la especie hexaploide F. arundinacea subsp. corsica parece ser un híbrido entre el diploide F. pratensis y el tetraploide F. arundinacea subsp. fenas. Se discuten diversas posibilidades sobre el origen del decaploide F. arundinacea var. letournexiana. Este trabajo contribuye a conocer mejor la filogenia de las festucas de hoja ancha y proporciona nueva información sobre
los cariotipos de estas especies —números cromosomáticos, niveles de ploidía y número y posición de loci de rDNA— mediante
FISH y la estima de sus tamaños genómicos mediante citometría de flujo.
Palabras clave: Festuca subgen. Schedonorus,
festucas de hoja ancha,
FISH,
tamaño genómico,
rADN.
|
INTRODUCTIONTOP
Subtribe Loliinae Dumort. belongs to the broad subfamily of grasses Pooideae Benth. —Poaceae Barnhart— and is formed by the
genus Festuca L. and 10 closely related genera (Catalán, 2006; Inda & al., 2008). This large group of temperate grasses is found over most of the world —except the Antarctic region—, with Eurasia being
the primary center of diversification. Festuca was morphologically divided into 2 main groups characterized according to the width of the leaves: the so-called “broad-leaved”
and “fine-leaved” fescues (Hackel, 1882). This separation has been supported by recent molecular phylogenetic analyses (Torrecilla & Catalán, 2002; Catalán & al., 2004; Inda & al., 2008).
The “Schedonorus -Lolium complex” is one of the main groups within the broad-leaved fescues. This complex comprises representatives of F. subgen. Schedonorus (P. Beauv) Petern., Lolium L., and Micropyropsis Romero Zarco & Cabezudo. The complex can be divided into two clades, the “European clade” and the “Maghrebian clade”, based
on the geographical location of most of their respective representatives. The divergence of these two clades has been supported
by molecular studies (Inda & al., 2014).
The area of origination of the Maghrebian clade was inferred to be the western Mediterranean region, where successive divergence
occurred during the early Pliocene and Pleistocene. This results in the dispersal of the clade mostly in NW Africa and SW
Europe (Inda & al., 2014). The speciation of the members of this clade was presumably associated with interspecific hybridization and several steps
of polyploidization (Borrill & al., 1977; Inda & al., 2014).
The Maghrebian clade involves only polyploid species, such as F. arundinacea subsp. fenas (Lag.) Bornm. —2n = 4x = 28—, F. mairei St.-Yves —2n = 4x = 28—, F. arundinacea subsp. corsica (Hack.) Kerguélen —2n = 6x = 42—, F. arundinacea subsp. atlantigena (St.-Yves) Auquier —2n = 8x = 56—, and F. arundinacea var. letourneuxiana (St.-Yves) Torrec. & Catalán —2n = 10x = 70—. On the other hand, the European clade is represented by both diploid —F. pratensis Huds., F. fontqueri St.-Yves, and Micropyropsis tuberosa Romero Zarco & Cabezudo— and polyploid species —tetraploid F. pratensis subsp. apennina De Not. and hexaploids F. arundinacea Schreb. subsp. arundinacea and F. gigantea (L) Vill.— (Borrill & al., 1977; Catalán & al., 2004; Hand & al., 2010; Inda & al., 2008, 2014).
The advent of molecular cytogenetics enabled to employ techniques such as fluorescent in situ hybridization —FISH— and genomic in situ hybridization —GISH— for phylogenetic studies. Humphreys & al. (1995) used GISH to indicate the origin of hexaploid F. arundinacea by interspecific hybridization of diploid F. pratensis and tetraploid F. arundinacea var. glaucescens Boiss. (F. arundinacea susbp. fenas). This hybridization was followed by whole genome duplication or alternatively, merge of unreduced gametes took place. Similarly,
GISH results confirmed that diploid F. pratensis is one of the progenitors of tetraploid F. pratensis subsp. apennina (Kopecký & al., unpubl.). This method has been also successfully used for the genome composition analysis of various interspecific
hybrids including ×Festulolium Asch. & Graebn. (Thomas & al., 1994; Kopecký & al., 2005). On the other hand, it was not possible to distinguish parental genomes in hybrids of Lolium perenne L. and Lolium multiflorum Lam. (Kopecký & al., 2008a), as well as progenitors of polyploid fine-leaved fescues from the Iberian Peninsula including F. ampla Hack. subsp. ampla, F. brigantina (Markgr.-Dann.) Markgr.-Dann., F. summilusitana Franco & Rocha Afonso, and F. duriotagana Franco & Rocha Afonso (Loureiro & al., 1997). Similarly, FISH with probes for specific DNA sequences can be employed for
evolutionary studies. The most frequently used are probes specific for 5S and 35S rDNAs. The number and position of rDNA loci
have been applied in many cytomolecular evolutionary analyses including the Schedonorus-Lolium complex (Thomas & al., 1996, 1997; Harper & al., 2004; Lideikyte, 2008; Inda & Wolny, 2013). Besides in situ hybridization techniques and many genetic markers, the estimations of genome size have also been used supportive tool to
indicate possible progenitors of allopolyploids and identify interspecific hybrids, where genome size of parents differ for
at least 15%, such as hybrids of Lolium multiflorum and F. pratensis (Huska & al., 2016; Kopecký & al., unpubl.). The advantage of flow cytometry is the relative low-cost and high-throughput for screening thousands
of such potential hybrids and allopolyploids.
Several species of F. subgen. Schedonorus, including tall fescue (F. arundinacea) and meadow fescue (F. pratensis), are widely used for forage and turf. Moreover, the ability of interspecific hybridization within Schedonorus-Lolium complex is recently used in grass breeding and several interspecific and intergeneric hybrids including hybrid ryegrass and
×Festulolium —hybrids of Festuca and Lolium— have been released and became popular among farmers (Kopecký & al., 2005, 2008a). Climatic changes call for implementation of new alleles into existing crop cultivars and thus, we can predict increasing
interest in the employment of other wild species in breeding. Despite the importance of this complex, only little is known
on the origination and the genomic composition of F. subgen. Schedonorus species, especially those of the Maghrebian clade. Such knowledge would be important for the targeted selection for interspecific
crosses.
The aim of our study was to determine the potential progenitor genomes of polyploid fescue species of the Maghrebian clade
using fluorescent in situ hybridization with probes specific for ribosomal DNAs —5S and 35S— and the estimation of genome size using flow cytometry
and with respect to the phylogenetic framework of this group obtained in our previous studies.
MATERIAL AND METHODSTOP
Plant materialTOP
We focused on various fescues from the Maghrebian clade. The seeds of these species were obtained from the germplasm banks
Western Regional Plant Introduction Station —WRPIS—, the Agriculture Service of the United States —USDA, USA—, and Aberystwyth
Genetic Resources —ABY, Aberystwyth, United Kingdom—. The species with their accessions are (Table 1): F. mairei: WRPIS PI-610941 —Morocco— and WRPIS PI-283312 —Morocco, cultivated in Sweden—; F. arundinacea subsp. fenas: WRPIS PI-289654 —Spain— and ABY-BN 354 —France, Rouen—; F. arundinacea subsp. corsica: ABY-BN 1175 —cultivated in Belgium, Liege—; F. arundinacea subsp. atlantigena: ABY-BN 865 and ABY-BN 807 —Morocco—; F. arundinacea var. letourneuxiana: ABY-BN 275 —Morocco— and ABY-BN 400 —Morocco.
Table 1. Summary of the number of chromosomes (2n), holoploid and monoploid genome size, and the numbers of 5S and 35S rDNA loci in various species of Festuca subgen. Schedonorus [a, Kopecký & al. (2010); b, Thomas & al. (1997), Lideikyte & al. (2008), and Ksiazczyk & al. (2010)].
| |
Chromosome number and ploidy level |
1C (pg) |
1Cx (pg) |
No. of 5S rDNA loci |
No. of 35S rDNA loci |
| F. pratensis |
2n = 2x = 14
|
3.25 ± 0.04a
|
3.25 |
2b
|
2b
|
| F. mairei (Morocco WRPIS PI-610941 & WRPIS PI-283312) |
2n = 4x = 28
|
5.02 ± 0.08 |
2.51 |
4 |
6 |
| F. arundinacea subsp. fenas (Spain WRPIS PI-289654 and France ABY-BN 354) |
2n = 4x = 28
|
5.24 ± 0.07 |
2.62 |
4 |
6 |
| F. arundinacea subsp. corsica (ABY-BN1175) |
2n = 6x = 42
|
8.31 ± 0.09 |
2.77 |
6 |
6 |
| F. arundinacea |
2n = 6x = 42
|
8.73 ± 0.04a
|
2.91 |
6b
|
4b
|
| F. arundinacea subsp. atlantigena (unknown origin ABY-BN 865 and Morocco ABY-BN 807)
|
2n = 8x = 56
|
8.11 ± 0.15 |
2.02 |
12 |
10 |
| F. arundinacea var. letourneuxiana (Morocco ABY-BN 275 & ABY-BN 400)
|
2n = 10x = 70
|
9.85 ± 0.04 |
1.97 |
16 |
12 |
Chromosome preparationsTOP
Caryopses samples of the cultivars were germinated in Petri dishes on wet filter paper and seedlings were planted in pots
in the greenhouse. Mitotic metaphase spreads have been prepared from root tips according to Jenkins & Hasterok (2007) and Valladolid & al. (2004). The caryopses without paleas and lemmas were germinated on filter paper moistened with water for 4 days at 20-22 °C in darkness.
Once geminated, the seeds were immersed in ice cold water for 24 h. They were then fixed in a mixture of methanol and glacial
acetic acid 3:1 —v/v— for 4 h at room temperature and stored at -20 °C until use. The root tips were cut —1 cm— and washed
several times with 0.01 M citrate buffer —pH 4.8—. The cell wall was digested by incubating the samples in an enzyme mixture
—4% pectinase from Aspergillus niger, 1% cellulase (Onozuka R-10), and 1% cellulase from Trichoderma viride in 10 mM citrate buffer pH 4.8— for 2 h at 37 °C. Thereafter, the squashing of root meristem tissue has been done in a drop
of 45% acetic acid. The slides were kept on dry ice for at least 30 min. The coverslips were then removed and the mixture
of absolute ethanol and glacial acetic acid —3:1 by volume— was poured over the slides followed by the immersion of the slides
in absolute ethanol for 30 min.
Fluorescent in situ hybridization (FISH) with rDNA probesTOP
FISH has been done according to Jenkins and Hasterok (2007) with minor modifications. DNA clone pTa794 containing 5S rDNA from Triticum aestivum L. was labelled by Nick translation with digoxigenin-dUTP —Roche— and the clone containing 35S rDNA region from Arabidopsis thaliana L. was labelled by Nick translation with tetramethyl-rhodamine-dUTP —Roche.
The slides were pretreated with RNase A —100 mg RNAse A, Sigma R-5503/ml of 10 mM Tris-HCl, and 15 mM NaCl— added to each
slide for 75 min/37 °C in humid chamber followed by the fixation of the slides with 1% formaldehyde in PBS —Sodium Phosphate
Buffered Saline pH 7— for 10 min/RT. The hybridization mixture involves 20 μl 100% deionized formamide, 8 μl 50% dextran sulfate
—w/v—, 4 μl 20x SSC, 2 μl 10% SDS —w/v—, 1 μl salmon sperm DNA, 2.5 μl 5S rDNA probe, and 2.5 μl 35S rDNA probe. Hybridization
mixture was denatured at 75 °C for 10 min. Thereafter, the mixture has been placed on slide with chromosome squashes and covered
with coverslip. The slides were denatured at 72 °C for 4:30 min and were kept in humid chamber for hybridization —16 h/37
°C—. After stringency washing, the sites of hybridization have been detected by the Anti-DIG-FITC conjugate —Roche—. Chromosomes
were counterstained with 0.2 mg/ml 4’,6-diamidino-2-phenylindole —DAPI— in the Vectashield antifade solution —Vector Laboratories—.
Slides were evaluated with Motic BA410 fluorescent microscope. Micrografx Picture Publisher and Adobe Photoshop software were
used for processing color pictures.
Holoploid genome size estimation using flow cytometryTOP
The leaves of adult plants growing in pots were used for flow cytometry analyses of genome size estimation. Nuclear suspensions
were prepared from 200 mg leaves of Festuca and 200 mg leaves of internal standard. We used either Pisum sativum L. ‘Ctirad’ —1C = 4.55 pg of DNA— or Secale cereale L. ‘Dankovske’ —1C = 8.09 pg DNA— as standards (Dolezel & al., 1998). Leaves in 500 μl Otto I reagent (Otto, 1992) were chopped by razor blade on a Petri dish. The suspension was filtered using 50 μm pore nylon filters and 1000 μl of Otto
II reagent —with propidium iodide— was added. Samples were analyzed using a CyFlow Space SYSMEX. At least 5,000 nuclei were
analyzed per sample. Each sample was analyzed three times during different days. Only measurements with coefficient of variation
—CV— below 3.5% have been considered in our study. 5 plants from each population were analyzed.
RESULTS AND DISCUSSIONTOP
Cytogenetic observations using FISH with rDNA probesTOP
Chromosome number has been determined for all the genotypes (Table 1). We confirmed tetraploidy in F. mairei and F. arundinacea subsp. fenas —2n = 4x = 28—, octoploidy in F. arundinacea subsp. atlantigena —2n = 8x = 56—, and decaploidy in F. arundinacea var. letourneuxiana —2n = 10x = 70—. We detected hexaploidy —2n = 6x = 42— in the accession ABY-BN 1175 of F. arundinacea subsp. corsica.
FISH resulted in the detection of 4 5S rDNA loci located proximally and 6 35S rDNA loci present in telomeric or subtelomeric
chromosome regions in both populations of Festuca arundinacea subsp. fenas —2n = 4x = 28—. Not a single chromosome carried both 5S and 35S rDNA loci (Fig. 1a). 4 5S rDNA and 6 35S rDNA loci have been detected in a Moroccan population of F. mairei —2n = 4x = 28—, all being present on different chromosomes (Fig. 1b). The other population, PI-283312, showed slightly different results. We detected the same number of 5S and 35S rDNA loci,
but one chromosome carried 2 signals of 35SrDNA in telomeric regions of both chromosome arms indicating non-homologous recombination
(Fig. 1c, marked by an arrow). Our results correlated with those of Thomas & al. (1997). They revealed that F. mairei and F. arundinacea subsp. glaucescens (F. arundinacea subsp. fenas) have the same numbers and pattern of 5S and 35S rDNA loci. However, we found the variation in the distribution of 35S rDNA
loci between 2 populations of F. mairei. This was not surprising, as the variation in the number of rDNA loci has been also reported in the other members of the
complex, like F. pratensis and Lolium perenne by Ksiazczyk & al. (2010).
|
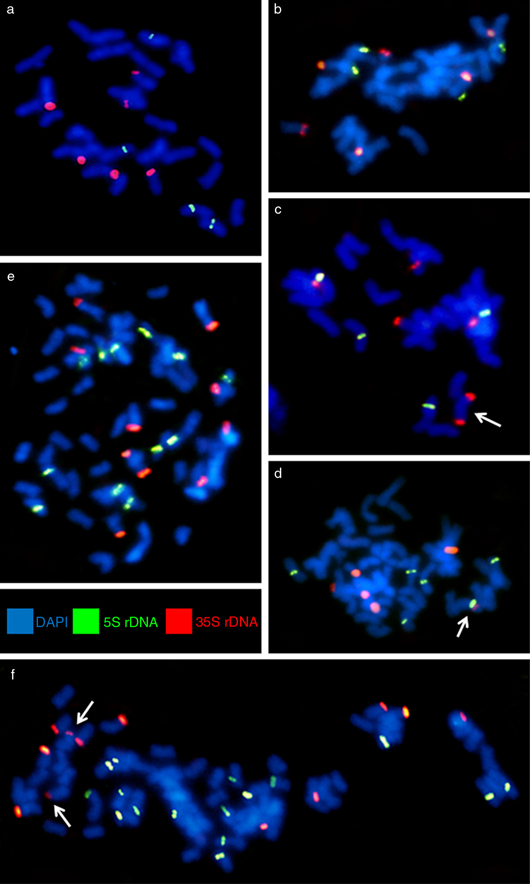 Fig. 1. Cytogenetic analysis of fescue species; FISH on metaphase plates of: a, Festuca arundinacea subsp. fenas; b, c, F. mairei; d, F. arundinacea subsp. corsica; e, F. arundinacea subsp. atlantigena; f, F. arundinacea var. letourneuxiana. [Probes for 35S rDNA in red color and for 5S rDNA in green color; chromosomes were counterstained using DAPI (blue color).] Fig. 1. Cytogenetic analysis of fescue species; FISH on metaphase plates of: a, Festuca arundinacea subsp. fenas; b, c, F. mairei; d, F. arundinacea subsp. corsica; e, F. arundinacea subsp. atlantigena; f, F. arundinacea var. letourneuxiana. [Probes for 35S rDNA in red color and for 5S rDNA in green color; chromosomes were counterstained using DAPI (blue color).]
|
|
The distribution and pattern of rDNAs in hexaploid F. arundinacea subsp. corsica, octoploid F. arundinacea subsp. atlantigena, and decaploid F. arundinacea var. letourneuxiana have been identified for the first time. In F. arundinacea subsp. corsica —2n = 6x = 42—, 6 5S rDNA loci and 6 loci of 35S rDNA have been detected (Fig. 1d). Surprisingly, one chromosome carried both 5S and 35S rDNA loci (Fig. 1d, indicated by an arrow), located close to each other. All 5S rDNA loci have been found in pericentromeric regions. In the closely related hexaploid F. arundinacea subsp. arundinacea, Thomas & al. (1997) confirmed F. pratensis and F. arundinacea subsp. glaucescens as progenitors of the hexaploid F. arundinacea subsp. arundinacea based on the number and position of rDNA loci. However, the authors found differences in the number and distribution of rDNA loci in F. arundinacea subsp. glaucescens and in the genome of F. arundinacea subsp. glaucescens that are present in the hexaploid F. arundinacea subsp. arundinacea. This suggests some genome modifications either in F. arundinacea subsp. glaucescens or in F. arundinacea subsp. arundinacea. Loss of rDNA loci inherited from diploid progenitors was reported in other allopolyploids (Vaughan & al., 1993; Leggett & Markhand, 1995).
We detected 12 loci of 5S rDNA and 10 loci of 35S rDNA in both populations of F. arundinacea subsp. atlantigena —2n = 8x = 56—. All the loci were detected on different chromosomes (Fig. 1e). 35S rDNA loci were located distally and 5S rDNA loci have been found in centromeric or pericentromeric regions. Similarly,
no variation in the numbers and positions of rDNA loci has been detected between 2 populations of decaploid F. arundinacea var. letourneuxiana —2n = 10x = 70—. Plants from both populations carried 16 loci of 5S rDNA preferentially located in proximal chromosome regions (Fig. 1f). In total, 12 loci of 35S rDNA, of which 2 are minor loci —indicated by arrows—, have been detected in telomeric or subtelomeric
regions. All rDNA loci were present on different chromosomes.
Genome size estimationsTOP
The estimation of genome size by flow cytometry revealed consistent results among populations of individual species (Table 1). The holoploid genome sizes —1C— in tetraploids ranged from 4.98-5.06 pg in F. mairei to 5.20-5.28 pg in F. arundinacea subsp. fenas. In F. arundinacea subsp. atlantigena and F. arundinacea var. letourneuxiana, the holoploid genome sizes were estimated to be 8.07-8.14 pg (Fig. 2a) and 9.85 pg, respectively. Hexaploid F. arundinacea subsp. corsica had the holoploid genome 1C = 8.31 pg (Fig. 2b).
|
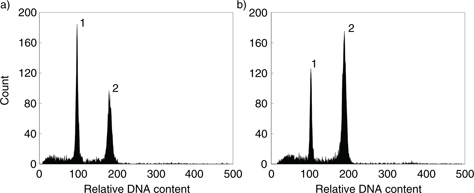 Fig. 2. Genome size estimation of fescue species using flow cytometry; histogram of relative fluorescence intensity obtained after simultaneous analysis of nuclei isolated from: a, Pisum sativum ‘Ctirad’ (1C = 4.55 pg, as an internal reference standard; G0/G1 nuclei, peak 1) and F. arundinacea subsp. atlantigena (G0/G1 nuclei, peak 2); b, Pisum sativum ‘Ctirad’ (1C = 4.55 pg, as an internal reference standard; G0/G1 nuclei, peak 1) and F. arundinacea subsp. corsica ABY-BN1175 (G0/G1 nuclei, peak 2). Fig. 2. Genome size estimation of fescue species using flow cytometry; histogram of relative fluorescence intensity obtained after simultaneous analysis of nuclei isolated from: a, Pisum sativum ‘Ctirad’ (1C = 4.55 pg, as an internal reference standard; G0/G1 nuclei, peak 1) and F. arundinacea subsp. atlantigena (G0/G1 nuclei, peak 2); b, Pisum sativum ‘Ctirad’ (1C = 4.55 pg, as an internal reference standard; G0/G1 nuclei, peak 1) and F. arundinacea subsp. corsica ABY-BN1175 (G0/G1 nuclei, peak 2).
|
|
Monoploid genome size —1Cx— decreases with increasing ploidy level (Table 1). Tetraploid subspecies —F. mairei and F. arundinacea subsp. fenas— displayed a monoploid genome size in range of 2.5-2.7 pg. Hexaploid population of F. arundinacea subsp. corsica has similar monoploid genome size as tetraploids —1Cx = 2.7 pg—. Monoploid genome size of octoploid F. arundinacea subsp. atlantigena has been estimated to 2.0 pg and in decaploid F. arundinacea var. letourneuxiana, 1Cx = 1.9 pg has been estimated.
The genome size of an allopolyploid could be expected as the sum of the genome sizes of its parents. This has been found in allopolyploids of the genera Triticum L., Arachis L., Glycine Willd., and Allium L. (Rees & Walters, 1965; Hammatt & al., 1991; Sing & al., 1996; Ohri & al., 1998). However, genome downsizing following polyploidy has been detected in other species including Brassica L. and Nicotiana L. allopolyploids (Ozkan & al., 2003). The loss of DNA after polyploidization has been frequently evidenced (Leitch & Bennett, 2004). Thus, the genome downsizing evidenced in our study is not exceptional (Fig. 3). Loureiro & al. (2007) also found a regression in monoploid genome size with increasing ploidy level in fine-leaved fescues. Highly polyploid species revealed significantly lower monoploid genome size compare to the diploids and tetraploids. Similarly, Šmarda & al. (2008) reported that hexaploid F. arundinacea subsp. arundinacea underwent the reduction of its genome after speciation from the cross of F. pratensis and F. arundinacea subsp. glaucescens. Several mechanisms including the elimination of non-coding DNA sequences during polyploid formation and the massive karyotype reconstruction including the deletions and non-homologous recombinations in the first generations are probably key factors in genome size reduction (Shaked & al., 2001).
|
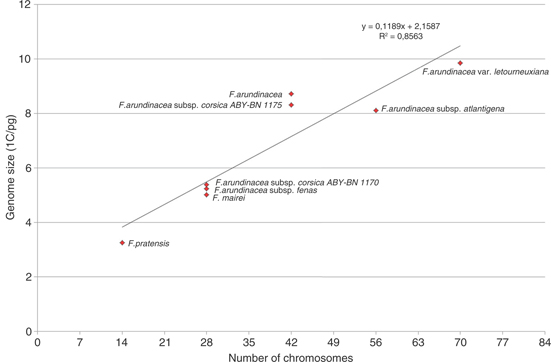 Fig. 3. Linear regression analysis between mean nuclear DNA content and chromosome number for various species of Festuca subgen. Schedonorus. Fig. 3. Linear regression analysis between mean nuclear DNA content and chromosome number for various species of Festuca subgen. Schedonorus.
|
|
In search of the ancestry of polyploid speciesTOP
There are only two tetraploids among Maghrebian broad-leaved fescues, F. arundinacea subsp. fenas (F. arundinacea subsp. glaucescens) and F. mairei. Using FISH, we revealed that both species have the same numbers and similar positions of rDNA loci and thus, are probably closely related. It may indicate that both species originated from the cross of the same parents. This is consistent with previous reports (Thomas & al., 1997; Catalán & al., 2004; Inda & al. 2008). Based on the karyotypic analysis and the presence and distribution of secondary constrictions, Malik & Thomas (1966) suggested that F. mairei and F. arundinacea subsp. glaucescens share one common genome. This has been supported by molecular sequence data of chloroplast and mitochondrial DNA and ITS
by Hand & al. (2010). Despite the close relationship of these species revealed by our FISH results supported by the phylogenetic study of Hand & al. (2010), we found difference in genome sizes of both species. This may indicate that they differ by at least one parental species.
Alternatively, they share the same progenitors, but massive genome reconstruction of one of the species took place during
its evolution, presumably genome downsizing of F. mairei.
Our FISH analysis of F. arundinacea subsp. corsica revealed that this species has highly similar number and position of rDNA loci as another hexaploid, F. arundinacea subsp. arundinacea from the European clade. This indicates that both hexaploids may have the same progenitors. Molecular analysis of ITS and GISH revealed that F. arundinacea subsp. arundinacea originated from the cross of maternal F. arundinacea subsp. glaucescens and paternal F. pratensis (Humphreys & al., 1994; Charmet & al., 1997). Similarly, phylogenetic analysis using sequence data of ITS and plastid trnT-L and trnL-F regions placed F. arundinacea subsp. corsica in the Maghrebian clade and indicated its close relationship with F. arundinacea subsp. fenas (Inda & al., 2014). Thus the later taxon could be one of the progenitors of F. arundinacea subsp. corsica. Based on the numbers and positions of rDNA loci revealed in this study and by Thomas & al. (1997), F. pratensis could be the second progenitor of F. arundinacea subsp. corsica. On the other hand, both species significantly differ in genome size —8.73 pg/1C in F. arundinacea vs. 8.31 pg/1C in F. arundinacea subsp. corsica; see Kopecký & al. (2010) and this study— and thus, F. arundinacea subsp. corsica genome probably underwent partial downsizing in larger extent than that assumed for F. arundinacea.
Apart from genome size changes, chromosome reshuffling probably took place after the speciation of both taxa. Non-reciprocal translocation indicated by the presence of single chromosome carrying both 5S and 35S rDNA loci has been found in F. arundinacea subsp. corsica (Fig. 1d). Similarly, structural changes of chromosomes after allopolyploidization have been detected in the hexaploid F. arundinacea subsp. arundinacea. Thomas & al. (1997) reported 4 loci of 35S rDNA in the hexaploid F. arundinacea, 2 of which belong to F. pratensis and 2 to F. arundinacea subsp. glaucescens. However, they found 4 loci of 35S rDNA in F. arundinacea subsp. glaucescens, and thus, 2 loci have been lost during the evolution of the hexaploid F. arundinacea subsp. arundinacea.
The origin and genome constitution of the octoploid F. arundinacea subsp. atlantigena is unclear. This species is closely related to F. mairei and also to F. arundinacea subsp. fenas. Thus it is possible that F. arundinacea subsp. atlantigena could have evolved by interspecific hybridization of these species either followed by whole genome duplication or alternatively, using unreduced gametes. This hypothesis is supported by the development of artificial hybrids of F. mairei and F. arundinacea subsp. fenas, which were fertile and morphologically close to F. arundinacea subsp. atlantigena (Chandrasekharan & Thomas, 1971). Similarly, Charmet & al. (1997) proposed F. mairei and F. arundinacea subsp. glaucescens as progenitors of F. arundinacea subsp. atlantigena and F. arundinacea var. letourneuxiana.
However, numbers of rDNA loci found in F. arundinacea subsp. atlantigena in this study are not a sum of rDNA loci detected in F. mairei and F. arundinacea subsp. fenas (Thomas & al., 1997). Thus, if these 2 species are really progenitors of F. arundinacea subsp. atlantigena, chromosome rearrangements including deletion of 2 35S rDNA loci and amplification/duplication of 4 5S rDNA loci likely took place. Moreover, the sum of genome sizes of both species highly exceeds the genome size of octoploid. Thus, if hypothesis on the origin of F. arundinacea subsp. atlantigena from the cross of F. mairei and F. arundinacea subsp. fenas is valid, a massive genome downsizing had to occur during the evolution of this species.
There are several scenarios for the potential speciation of the decaploid F. arundinacea var. letourneuxiana. Borrill (1972) speculated that it originated from the cross of F. arundinacea subsp. arundinacea (6x) and F. mairei (4x) followed by whole genome duplication. However, positions and number of rDNA loci —16 and 12 loci of 5S and 35S rDNA, respectively— in F. arundinacea var. letourneuxiana are highly different from the sum of both potential parents. Moreover, this scenario requires downsizing for over 30% of the genome after potential allopolyploidization, based on our genome size estimations.
Alternatively, F. arundinacea var. letourneuxiana could be a product of interspecific hybridization of the octoploid F. arundinacea subsp. atlantigena and the diploid F. pratensis followed by whole genome duplication. In this case, only the amplification of 2 5S rDNA loci would be necessary with only minor reduction of genome size. However, F. pratensis possesses 2 loci of 35S rDNA located proximally in the short arm of chromosome 3F (Kopecký & al., 2008b) and we did not identify such positioning of 35S rDNA in F. arundinacea var. letourneuxiana in this study.
The close phylogenetic relationship of F. arundinacea subsp. atlantigena and F. arundinacea var. letourneuxiana indicates that the former one has been involved in the origination of the latter. Moreover, the molecular analyses of cpDNA and mtDNA sequences suggest that F. arundinacea subsp. atlantigena serves as maternal progenitor (Hand & al., 2010; Inda & al., 2014). Based on the above mentioned findings and our FISH and genome size results, we hypothesize that this decaploid originated from the merge of an unreduced gamete —egg cell— of F. arundinacea subsp. atlantigena with a reduced gamete —pollen— of either F. mairei or F. arundinacea subsp. fenas, more presumably the former one. Such scenario would require amplification/duplication of 2 5S rDNA loci and the loss of 1 —odd— 35S rDNA loci. The sum of both potential progenitor genome sizes is almost equal to the genome size estimated for F. arundinacea var. letourneuxiana —10.62pg/1C vs. 9.85pg/1C—. Alternatively, a currently unknown or extinct diploid parent could be the missing 2x parent of the decaploid F. arundinacea var. letourneuxiana.
Concluding remarksTOP
Our study provided new information on the genome size and the numbers and distributions of rDNA loci of broad-leaved fescues
from Maghrebian clade and improved the knowledge on the phylogeny of this group of grasses —a hypothetical scenario on the
evolution of the group is summarized in Fig. 4—. We found that F. arundinacea subsp. corsica presumably resulted from the interspecific hybridization of the diploid F. pratensis and the tetraploid F. arundinacea subsp. fenas and thus, has probably the same progenitors as F. arundinacea subsp. arundinacea from the European clade. The origin of the octoploid F. arundinacea subsp. atlantigena remains unclear, however, it probably originated from crossing of the tetraploids Festuca arundinacea subsp. fenas —2n = 4x = 28— and Festuca mairei —2n = 4x = 28— followed by whole genome duplication and massive genome reconstruction. The most probable scenario on the origin of
the decaploid F. arundinacea var. letourneuxiana involves merge of an unreduced gamete of the octoploid F. arundinacea subsp. atlantigena and a reduced gamete of tetraploid fescue, presumably F. mairei.
|
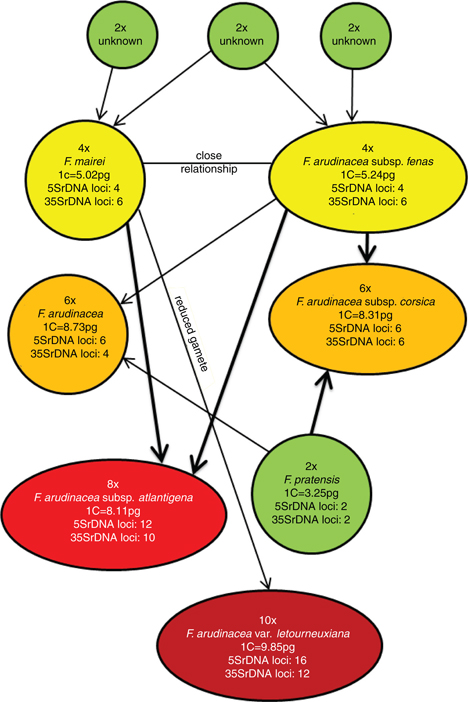 Fig. 4. Hypothetical scenario on the origin of polyploid fescues from Marghrebian clade. Fig. 4. Hypothetical scenario on the origin of polyploid fescues from Marghrebian clade.
|
|
ACKNOWLEDGEMENTSTOP
We are grateful to Dr. Betehktin and Prof. R. Hasterok (University of Silesia, Katowice) for providing the 5S and 35S clones,
the seed banks of Western Regional Plant Introduction Station (WRPS), Agriculture Service of the United States (Pullman, USA),
and Aberystwyth Genetic Resources (ABY, Ian Thomas) (Aberystwyth, United Kingdom) for seeds of analyzed species and Dr. P.
Catalán for critical reading and valuable comments. The study was funded by the Spanish Ministry of Science (grant n.º CGL2012-39953-C02-01)
and Aragon Government and European Social Fund grant to our Bioflora research group.
REFERENCESTOP
| ○ |
Borrill, M. 1972. Studies in Festuca III. The contribution of F. scariosa to the evolution of polyploids in sections Bovinae and Scariosae. New Phytologist 71: 523-532. http://dx.doi.org/10.1111/j.1469-8137.1972.tb01953.x.
|
| ○ |
Borrill, M., Kirby, M. & Morgan, W.G. 1977. Studies in Festuca 11. Interrelationships of some putative diploid ancestors of the polyploid broad-leaved fescues. New Phytologist 78: 661-674. http://dx.doi.org/10.1111/j.1469-8137.1977.tb02171.x.
|
| ○ |
Catalán, P., Torrecilla, P., Rodríguez, J.A.L. & Olmstead, R.G. 2004. Phylogeny of the festucoid grasses of subtribe Loliinae and allies (Poeae, Pooideae) inferred from ITS and trnL-F sequences. Molecular Phylogenetics and Evolution 31: 517-541. http://dx.doi.org/10.1016/j.ympev.2003.08.025.
|
| ○ |
Catalán, P. 2006. Phylogeny and evolution of Festuca L. and related genera of subtribe Loliinae (Poeae, Poaceae). In: Sharma, A.K. & Sharma, A. (eds.), Plant Genome. Biodiversity and Evolution. Pp. 255-303. Enfield: Science Publishers.
|
| ○ |
Chandrasekharan, P. & Thomas, H. 1971. Studies in Festuca. V. Cytogenetic relationships between species of Bovinae and Scariosae. Zeitschrift für Pflanzenzuchtung 65: 353-354.
|
| ○ |
Charmet, G., Ravel, C. & Balfourier, F. 1997. Phylogenetic analysis in the Festuca-Lolium complex using molecular markers and ITS rDNA. Theoretical and Applied Genetics 94: 1038-1046. http://dx.doi.org/10.1007/s001220050512.
|
| ○ |
Dolezel, J., Greilhuber, J., Lucretti, S., Meister, A., Lysak, M., Nardi, L. & Obermayer, R. 1998. Plant genome size estimation by flow cytometry: interlaboratory comparison. Annals of Botany 82: 17-26.
|
| ○ |
Hackel, E. 1882. Monografia Festucarum Europearum. Kassel and Berlin: T. Fischer.
|
| ○ |
Hammatt, N., Blackall, N.W. & Davey, M.R. 1991. Variation in the DNA content of Glycine species. Journal of Experimental Botany 42: 659-665. http://dx.doi.org/10.1093/jxb/42.5.659.
|
| ○ |
Hand, M., Cogan, N.O.I., Stewart, A.V. & Forster, J.W. 2010. Evolutionary history of tall fescue morphotypes inferred from molecular phylogenetics of the Lolium-Festuca species complex. BMC Evolutionary Biology 10: 303-320. http://dx.doi.org/10.1186/1471-2148-10-303.
|
| ○ |
Harper, J.A., Thomas, I.D., Lovatt, J.A. & Thomas, H.M. 2004. Physical mapping of rDNA sites in possible diploid progenitors of polyploid Festuca species. Plant Systematics and Evolution 245: 163-168. http://dx.doi.org/10.1007/s00606-003-0110-2.
|
| ○ |
Humphreys, M.W., Thomas, H.M., Morgan, W.G., Meredith, M.R., Harper, J.A., Thomas, H., Zwierzykowski, Z. & Ghesquiére, M. 1995. Discriminating the ancestral progenitors of hexaploid Festuca arundinacea using genomic in situ hybridization. Heredity 75: 171-174. http://dx.doi.org/10.1038/hdy.1995.120.
|
| ○ |
Huska, D., Leitch, I.J., De Carvalho, J.F., Leitch, A.R., Salmon, A., Ainouche, M. & Kovarik, A. 2016. Persistence, dispersal and genetic evolution of recently formed Spartina homoploid hybrids and allopolyploids in Southern England. Biological Invasions 18: 2137-2151. http://dx.doi.org/10.1007/s10530-015-0956-6.
|
| ○ |
Inda, L.Á., Segarra-Moragues, J.G., Müller, J., Peterson, P.M. & Catalán, P. 2008. Dated historical biogeography of the temperate Loliinae (Poaceae, Pooideae) grasses in the northern and southern hemispheres. Molecular Phylogenetics and Evolution 46: 932-957. http://dx.doi.org/10.1016/j.ympev.2007.11.022.
|
| ○ |
Inda, L. Á., Sanmartín, I., Buerki, S. & Catalán, P. 2014. Mediterranean origin and Miocene-Holocene Old World diversification of meadow fescues and ryegrasses (Festuca subgenus Schedonorus and Lolium). Journal of Biogeography 41: 600-614. http://dx.doi.org/10.1111/jbi.1221.
|
| ○ |
Inda, L. Á. & Wolny, E. 2013. Fluorescent in situ hybridization of the ribosomal RNA genes (5S and 35S) in the genus Lolium: Lolium canariense, the missing link with Festuca? Anales del Jardín Botánico de Madrid 70: 97-102. http://dx.doi.org/10.3989/ajbm.2329.
|
| ○ |
Jenkins, G. & Hasterok, R. 2007. BAC ´landing´on chromosomes of Brachypodium distachyon for comparative genome alignment. Nature Protocols 2: 88-98. http://dx.doi.org/10.1038/nprot.2006.490.
|
| ○ |
Kopecký, D., Lukaszewski, A.J. & Doležel, J. 2005. Genomic constitution of Festulolium cultivars released in the Czech Republic. Plant Breeding 124: 454-458. http://dx.doi.org/10.1111/j.1439-0523.2005.01127.x.
|
| ○ |
Kopecký, D., Lukaszewski, A.J. & Doležel, J. 2008a. Cytogenetics of Festulolium (Festuca × Lolium hybrids). Cytogenetics and Genome Research 120: 370-383. http://dx.doi.org/10.1159/000121086.
|
| ○ |
Kopecký, D., Lukaszewsk, A.J. & Doležel, J. 2008b. Meiotic behavior of individual chromosomes of Festuca pratensis in tetraploid Lolium multiflorum. Chromosome Research 16: 987-998. http://dx.doi.org/10.1007/s10577-008-1256-0.
|
| ○ |
Kopecký, D., Havránková, M., Loureiro, J., Castro, S., Lukaszewski, A.J., Bartoš, J., Kopecká, J. & Doležel, J. 2010. Physical distribution of homoeologous recombination in individual chromosomes of Lolium multiflorum/Festuca pratensis substitutions. Cytogenetic and Genome Research 129: 162-172. http://dx.doi.org/10.1159/000313379.
|
| ○ |
Ksiazczyk, T., Taciak, M. & Zwierzykowski, Z. 2010. Variability of ribosomal DNA sites in Festuca pratensis, Lolium perenne, and their intergeneric hybrids, revealed by FISH and GISH. Journal of Applied Genetics 51: 449-60. http://dx.doi.org/10.1007/BF03208874.
|
| ○ |
Leggett, J.M. & Markhand, G.S. 1995. The genomic identification of some monosomics of Avena sativa L-Cv Sun-Ii using genomic in situ hybridization. Genome 38: 747-751. http://dx.doi.org/10.1139/g95-094.
|
| ○ |
Leitch, I.J. & Bennett, M.D. 2004. Genome downsizing in polyploid plants. Biological Journal of the Linnean Society 82: 651-663. http://dx.doi.org/10.1111/j.1095-8312.2004.00349.x.
|
| ○ |
Lideikyte, L., Pasakinskiene, I., Lemeziene, N., Nekrosas, S. & Kanapeckas, J. 2008. FISH assessment of ribososmal DNA sites in the chromosome sets of Lolium, Festuca and Festuloium. Zemdirbyste 95: 116-124.
|
| ○ |
Loureiro, J., Kopecký, D., Castro, S., Santos, C. & Silveira, P. 2007. Flow cytometric and cytogenetic analyses of Iberian Peninsula fescues. Plant Systematics and Evolution 269: 89-105. http://dx.doi.org/10.1007/s00606-007-0564-8.
|
| ○ |
Malik, C.P. & Thomas, P.T. 1966. Kariotypic studies in some Lolium and Festuca species. Caryologia 19: 167-196. http://dx.doi.org/10.1080/00087114.1966.10796216.
|
| ○ |
Ohri, D., Fritsch, R.M. & Hanelt, P. 1998. Evolution of genome size in Allium (Alliaceae). Plant Systematics and Evolution 210: 57-86. http://dx.doi.org/10.1007/BF00984728.
|
| ○ |
Otto, F. 1992. Preparation and staining of cells for high-resolution DNA analysis. In: Radbruch A. (ed.), Flow cytometry and cell sorting. Pp. 65-68. Berlin: Springer-Verlag. http://dx.doi.org/10.1007/978-3-662-02785-1_8.
|
| ○ |
Ozkan, H., Tuna, M. & Arumuganathan, K. 2003. Nonadditive Changes in genome Size During Allopolyploidization in the Wheat (Aegilops-Triticum) Group. Journal of Heredity 94: 260-264. http://dx.doi.org/10.1093/jhered/esg053.
|
| ○ |
Rees, H. & Walters, M.R. 1965. Nuclear DNA and the evolution of wheat. Heredity 20: 73-82. http://dx.doi.org/10.1038/hdy.1965.9.
|
| ○ |
Shaked, H., Kashkush, K., Ozkan, H., Feldman, M. & Levy, A.A. 2001. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat. Plant Cell 13: 1749-1759. http://dx.doi.org/10.1105/TPC.010083.
|
| ○ |
Sing, K.P., Raina, S.N. & Singh, A.K. 1996. Variation in chromosomal DNA associated with the evolution of Arachis species. Genome 39: 890-897. http://dx.doi.org/10.1139/g96-112.
|
| ○ |
Šmarda, P., Bures, P., Horova, L., Foggi, B. & Rossi, G. 2008. Genomic Size and GC Content Evolution of Festuca: Ancestral Expansion and Subsequent Reduction. Annals of Botany 101: 421-433. http://dx.doi.org/10.1093/aob/mcm307.
|
| ○ |
Thomas, H.M., Harper, J.A., Meredith, M.R., Morgan, W.G. & King, I.P. 1997. Physical mapping of ribosomal DNA sites in Festuca arundinacea and related species by in situ hybridization. Genome 40: 406-410. http://dx.doi.org/10.1139/g97-054.
|
| ○ |
Thomas, H.M., Harper, J.A., Meredith, M.R., Morgan, W.G., Thomas, I.D., Timms, E. & King, I.P. 1996. Comparison of ribosomal DNA sites in Lolium species by fluorescence in situ hybridization. Chromosome Research 4: 486-490. http://dx.doi.org/10.1007/BF02261775.
|
| ○ |
Thomas, H.M., Morgan, W.G., Meredith, M.R., Humphreys, M.W., Thomas, H. & Leggett, J.M. 1994. Identification of parental and recombined chromosomes in hybrid derivatives of Lolium multiflorum x Festuca pratensis by genomic in situ hybridization. Theoretical and Applied Genetics 88: 909-13. http://dx.doi.org/10.1007/BF00220795.
|
| ○ |
Torrecilla, P. & Catalán, P. 2002. Phylogeny of Broad-leaved and Fine-leaved Festuca Lineages (Poaceae) based on Nuclear ITS Sequences. Systematic Botany 27: 241-251. http://dx.doi.org/10.1043/0363-6445-27.2.241.
|
| ○ |
Valladolid, A., Blas, R. & Gonzáles, R. 2004. Introducción al recuento de cromosomas somáticos en raíces andinas. In: Seminario, J. (ed.), Conservación y uso de la biodiversidad de raíces y tubérculos andinos: Una década de investigación para el desarrollo (1993-2003). Pp. 95-99. Lima: Centro Internacional de la Papa.
|
| ○ |
Vaughan, H.E., Jamilena, M., Rejon, C.R., Parker, J.S. & Garridoramos, M.A. 1993. Loss of nucleolar-organizer regions during polyploid evolution in Scilla autumnalis. Heredity 71: 574-580. http://dx.doi.org/10.1038/hdy.1993.181.
|